DOTTORATO DI RICERCA in biofarmaceutica e...
160
UNIVERSITÀ DEGLI STUDI DI PARMA ISTITUTO DI SCIENZE BIOCHIMICHE DOTTORATO DI RICERCA IN BIOFARMACEUTICA - FARMACOCINETICA VIII CICLO (1992-1995) FARMACOCINETICA DELLA VITAMINA A: CARATTERIZZAZIONE DELLA REAZIONE METABOLICA RESPONSABILE DELLA BIOSINTESI DI ACIDO RETINOICO Title: rea.ps Creator: RasMol Version 2.5 CreationDate: DAVIDE CAVAZZINI
-
Upload
nguyencong -
Category
Documents
-
view
215 -
download
0
Transcript of DOTTORATO DI RICERCA in biofarmaceutica e...

UNIVERSITÀ DEGLI STUDI DI PARMA
ISTITUTO DI SCIENZE BIOCHIMICHE
DOTTORATO DI RICERCA IN BIOFARMACEUTICA - FARMACOCINETICA
VIII CICLO (1992-1995)
FARMACOCINETICA DELLA VITAMINA A: CARATTERIZZAZIONE DELLA REAZIONE
METABOLICA RESPONSABILE DELLA BIOSINTESI DI ACIDO RETINOICO
Title: rea.psCreator: RasMol Version 2.5CreationDate:
DAVIDE CAVAZZINI

UNIVERSITÀ DEGLI STUDI DI PARMA
ISTITUTO DI SCIENZE BIOCHIMICHE
DOTTORATO DI RICERCA IN BIOFARMACEUTICA - FARMACOCINETICA
VIII CICLO (1992-1995)
FARMACOCINETICA DELLA VITAMINA A: CARATTERIZZAZIONE DELLA REAZIONE
METABOLICA RESPONSABILE DELLA BIOSINTESI DI ACIDO RETINOICO
DAVIDE CAVAZZINI

Indice
INDICE
1. SOMMARIO...........................................................................................1
2. INTRODUZIONE...................................................................................3
2.1 I RETINOIDI: CARATTERISTICHE FISICO-CHIMICHE ED EFFETTI BIOLOGICI.............32.11 Il retinolo...........................................................................................................32.12 La retinaldeide...................................................................................................42.13 L’acido retinoico...............................................................................................4
2.2 MECCANISMO D'AZIONE NUCLEARE DELL'ACIDO RETINOICO...............................6
2.3 I RETINOIDI COME FARMACI................................................................................9
2.4 ASSUNZIONE E TRASPORTO DI RETINOLO............................................................132.41 Assorbimento intestinale di retinoidi e metabolismo epatico..............................132.42 Trasporto plasmatico di retinolo: la RBP...........................................................152.43 Il rilascio del retinolo ai tessuti..........................................................................16
2.5 METABOLISMO INTRACELLULARE DEL RETINOLO...............................................162.51 I trasportatori intracellulari dei retinoidi..........................................................16 2.52 L’esterificazione del retinolo.............................................................................22 2.53 Idrolisi dei retinil esteri.....................................................................................23 2.54 Biosintesi dell’acido retinoico...........................................................................25 2.55 Il sistema “misto”..............................................................................................292.56 Il sistema “solubile”..........................................................................................30
3. FINALITÀ DELLA RICERCA.............................................................33
4. RISULTATI............................................................................................34
4.1 OTTIMIZZAZIONE DEL SAGGIO IN VITRO PER LA SINTESI DI ACIDO RETINOICO......34
4.2 IL SISTEMA “MISTO” IN ESTRATTI SUBCELLULARI DA FEGATO BOVINO..............................................................................................................36
I
Rossi Marco & Carlo, 03/01/-0001,

Indice
4.3 PARZIALE PURIFICAZIONE DELL’ATTIVITÀ ENZIMATICA SOLUBILE DA CITOSOL DI FEGATO BOVINO...............................................................................37
4.4 IDENTIFICAZIONE E CARATTERIZZAZIONE DELLE VIE DI BIOSINTESI DI RA IN ESTRATTI CELLULARI DA FEGATO DI RATTO. CONFRONTO CON L’ATTIVITÀ IDENTIFICATA IN ORGANI BOVINI......................................................40
4.5 IPOTESI SULLA FORMAZIONE DEL ACIDO 9-CIS RETINOICO..................................44
4.6 ISOMERIZZAZIONE DI ACIDO ALL-TRANS RETINOICO A ACIDO 9-CISRETINOICO..........................................................................................................45
4.7 ANALISI DELLA CONFIGURAZIONE ISOMERICA DELL’ACIDO RETINOICO SINTETIZZATO IN PRESENZA DEI SISTEMI “MISTO” E “SOLUBILE” A PARTIRE DA ALL-TRANS RETINOLO-CRBPI.........................................................48
4.8 IL COMPLESSO 9-CIS RETINOLO-CRBPI..............................................................49
4.9 STEREOSELETTIVITÀ DELLE VIE DI SINTESI “MISTA” E “SOLUBILE” NELLA FORMAZIONE DI RA DA ALL-TRANS RETINOLO IN FORMA LIBERA O LEGATA A CRBPI...............................................................................52
4.10 EFFETTO DELLA APOCRBPI SULLA SINTESI DI RA DA ALL-TRANS RETINOLO-CRBPI..............................................................................................54
4.11 STEREOSELETTIVITÀ DELLE VIE DI SINTESI “MISTA” E “SOLUBILE” PER LA FORMAZIONE DI RA DA 9-CIS RETINOLO IN FORMA LIBERA O LEGATA A CRBPI...............................................................................................56
4.12 DETERMINAZIONE DEI LIVELLI DI 9-CIS RETINOLO NELLA FRAZIONE MICROSOMALE E CITOSOLICA E SUO POSSIBILE RUOLO QUALE PRECURSORE DEL 9-CIS RA................................................................................62
4.13 PARZIALE PURIFICAZIONE E CARATTERIZZAZIONEDELL’ATTIVITÀ SOLUBILE DA FEGATO DI RATTO.................................................65
4.14 BIOSINTESI DI ACIDO RETINOICO IN CELLULE SOMATICHE DI TESTICOLO DI RATTO..........................................................................................69
5. DISCUSSIONE.......................................................................................77
6. CONCLUSIONI.....................................................................................88
7. MATERIALI E METODI...........................................................................90
II

Indice
7.1 CONSERVAZIONE E STABILITÀ DEI RETINOIDI......................................................90
7.2 ESPRESSIONE E PURIFICAZIONE DI CRBPI E CRBPII RICOMBINANTE DA ESCHERICHIA COLI IN FORMA APO E OLO.............................................................90
7.3 ESPRESSIONE E PURIFICAZIONE DI CRABPI RICOMBINANTE DA ESCHERICHIA COLI IN FORMA APO E OLO.............................................................92
7.4 STIMA DELLA CONCENTRAZIONE E DEL GRADO DI SATURAZIONE DELLE RETINOID-BINDING PROTEIN.......................................................................94
7.5 ELETTROFORESI DI CRBP E CRABP..................................................................94
7.6 RIMOZIONE DEL LIGANDO DA CRBP E CRABP..................................................95
7.7 RIMOZIONE DEI RETINOIDI PRESENTI IN FORMA LIBERA......................................95
7.8 FRAZIONAMENTO SUBCELLULARE......................................................................96
7.9 ANALISI PER SEDIMENTAZIONE IN GRADIENTE DI GLICEROLO.............................97
7.10 PARZIALE PURIFICAZIONE DELL’ATTIVITÀ RETINOICO BIOSINTETICA DA CITOSOL DI FEGATO DI RATTO............................................................................97
7.11 SAGGIO DI BIOSINTESI IN VITRO DI ACIDO RETINOICO.........................................97
7.12 ESTRAZIONE DEI RETINOIDI................................................................................98
7.13 ANALISI IN HPLC DEI RETINOIDI........................................................................100
7.14 PREPARAZIONE DI 9-CIS RETINOLO MEDIANTE RIDUZIONE DI 9-CIS RETINALDEIDE...........................................................................................101
7.15 MISURE DI FLUORESCENZA.................................................................................101
8 BIBLIOGRAFIA....................................................................................103
III

Indice
9 PUBBLICAZIONI..................................................................................109
IV

Sommario
1. SOMMARIO
I retinoidi sono costituiti da una classe di composti lipofili, naturali e di sintesi, che include il retinolo (Vitamina A) e altre sostanze strutturalmente e funzionalmente correlate. L'acido retinoico, metabolita naturale che si genera dall'ossidazione intracellulare del retinolo introdotto con la dieta, è il retinoide biologicamente più attivo: riveste un ruolo essenziale nella modulazione e il mantenimento di importanti processi cellulari, quali la proliferazione e il differenziamento di diversi tessuti, ed è essenziale per il processo di morfogenesi e lo sviluppo embrionale (Sporn, M.B., Roberts, A.B., and Goodman, D.S. Eds. (1994), The Retinoids: Biology, Chemistry and Medicine, Raven Press Ltd., New York).Oltre a far regredire, in vitro, il fenotipo neoplastico di numerose linee cellulari tumorali, l'acido retinoico è in grado, in vivo, di normalizzare numerosi disordini proliferativi e differenziativi dei tessuti epiteliali, essendo impiegato nel trattamento di varie forme di dermatosi (Es. acne nodulocistica, psoriasi, ittiosi), ed è stato utilizzato recentemente con successo nella terapia di pazienti con leucemia acuta promielocitica.Si ritiene che l'acido retinoico esplichi tali effetti con meccanismo analogo a quello degli ormoni steroidei: sono stati identificate due famiglie di recettori nucleari, denominati RAR (retinoic acid receptors) e RXR (retinoid X receptors), che modulano la trascrizione di specifici geni in seguito al legame rispettivamente con gli isomeri all-trans e 9-cis dell'acido retinoico.Diversi retinoidi, naturali e di sintesi, sono oggi utilizzati estensivamente in campo dermatologico, e nella prevenzione o trattamento di varie forme neoplastiche, spesso con ottimi risultati. Uno degli ostacoli maggiori all'impiego terapeutico dell'acido retinoico e dei suoi derivati di sintesi è costituito dal basso indica terapeutico: alle dosi comunemente impiegate essi provocano numerosi e a volte gravi effetti collaterali. Sono inoltre notevoli le limitazioni imposte dalla loro attività teratogena ed embriotossica.Tra i vari eventi farmacocineticamente significativi che portano i retinoidi, somministrati per via orale o topica, ad esplicare effetti pleiotropici a livello cellulare, notevoli progressi sono stati fatti, ad esempio, nella comprensione del meccanismo molecolare d'azione a livello nucleare, ma ancora relativamente oscuri sono i processi che riguardano la biotrasformazione del retinolo ad acido retinoico.Gli obiettivi primari del presente lavoro di ricerca sono stati quelli di chiarire alcuni aspetti di questa fase della farmacocinetica della Vitamina A, ritenendo che la conoscenza approfondita delle modalità con cui avviene la sintesi di acido retinoico sia di primaria importanza al fine di poter controllare e regolare le numerose ed essenziali attività biologiche delle quali tale molecola è dotata.I risultati ottenuti i hanno permesso di fare maggiore chiarezza sull’identità dei sistemi enzimatici e dei substrati fisiologici responsabili della biosintesi di acido retinoico, hanno
1

Sommario
portato alla parziale purificazione e caratterizzazione biochimica e molecolare di tali attività in differenti tessuti ed organi animali, e hanno identificato la forma isomerica dei prodotti sintetizzati in vitro.
2

Introduzione
2. INTRODUZIONE
2.1 I RETINOIDI: CARATTERISTICHE FISICO-CHIMICHE ED EFFETTI BIOLOGICI
Il termine “retinoidi” indica una famiglia di sostanze che include composti naturali quali la vitamina A (il retinolo) e i suoi metaboliti retinaldeide ed acido retinoico, oltre a molecole di sintesi con simili caratteristiche chimiche e attività biologica I retinoidi sono molecole a carattere lipofilo, estremamente sensibili se esposte alla luce, all'ossigeno o alle alte temperature, la cui struttura, in forma più o meno modificata, è essenzialmente costituita e da un anello trimetilcicloesenico e da una catena isoprenica contenente il gruppo funzionale (Figura 1) (Sporn, Roberts, e Goodman Eds. (1994), “The Retinoids: Biology, Chemistry and Medicine” Raven Press Ltd., New York).
Figura 1: Formule di struttura dei principali retinoidi
2.11 Retinolo
Il retinolo (Vitamina A alcool) rappresenta la principale forma nella quale i retinoidi sono assorbiti con la dieta e trasportati agli organi bersaglio. Esso costituisce il precursore per
3

Introduzione
la sintesi di retinaldeide e acido retinoico, (assunti in quantità trascurabile con la dieta), ed è l'unico retinoide a poter essere accumulato dai vari tessuti, sotto forma di retinil estere (Levin, M.S., 1994). Da tempo è stata riconosciuta l'importanza del retinolo (ROH) come molecola essenziale per il mantenimento di numerosi ed importanti processi biologici: esso costituisce il precursore dei pigmenti visivi, è un nutriente necessario per il mantenimento della crescita e il corretto differenziamento di tutti i tessuti epiteliali, svolge un ruolo importante nella regolazione della risposta del sistema immunitario, ed è inoltre indispensabile per lo sviluppo embrionale e la morfogenesi, oltre che per il mantenimento della spermatogenesi (Wolf, 1984). Il retinolo è la molecola che per prima è stata impiegata per il trattamento di alcune forme di dermatosi, oltre che nella prevenzione ed il trattamento di alcune neoplasie, ma il suo uso è stato limitato dalla scarsa efficacia terapeutica e dai notevoli effetti collaterali (ipervitaminosi A).
2.12 Retinaldeide
Essendo un importante componente funzionale dei fotorecettori, la retinaldeide (RALD) rappresenta la molecola chiave per il mantenimento della funzione visiva. Assunta con la dieta in quantità trascurabile, viene sintetizzata nei tessuti oculari in seguito all'ossidazione del retinolo (Wald, 1968). Molecola chimicamente instabile ed estremamente reattiva, la retinaldeide è praticamente assente nei tessuti extraoculari (Wolf, 1984), nei quali potrebbe però costituire l’intermedio transiente nella reazione metabolica di biosintesi di acido retinoico (Ottonello et al., 1994; Posch et al., 1992a)
2.13 Acido retinoico
L’acido retinoico (RA), il retinoide biologicamente più attivo e potente, molecola indispensabile per la corretta proliferazione ed il differenziamento in diversi tessuti embrionali ed adulti, si forma per ossidazione intracellulare del retinolo (Napoli, 1994).L’acido retinoico è essenziale nel modulare la crescita ed il differenziamento in tutti i tessuti epiteliali: è in grado di arrestare in vitro il processo di cheratinizzazione della trachea in colture d'organo, così come la formazione e la stratificazione dello strato corneo nei cheratinociti, nei quali inibisce la sintesi di vari tipi di cheratina (De Luca, 1991). Ma l’acido retinoico esercita un’importante azione regolatoria sulla proliferazione e sul differenziamento degli epiteli anche in vivo, tanto che frequente è l’impiego oggi di tale composto, o dei suoi analoghi naturali o di sintesi, per il trattamento di varie forme di dermatosi (pelle fotodanneggiata, acne, psoriasi, ittiosi) (Fritsch, 1992).I notevoli effetti esercitati dall’acido retinoico su diverse linee cellulari (tumorali) (Es. cellule di carcinoma embrionale F9, cellule leucemiche mieloidi, cellule di neuroblastoma...), nelle quali è in grado di revertire in modo stabile il fenotipo
4

Introduzione
neoplastico, sono studiati con grande attenzione, anche per le importanti applicazioni terapeutiche che ne potrebbero derivare (Smith et al., 1992).In queste colture cellulari l’acido retinoico stimola il processo differenziativo modulando la trascrizione di specifici geni: nel caso delle cellule di carcinoma embrionale, ad esempio, l’espressione delle proteine laminina B1, fosfatasi alcalina, e collagene è indotta poche ore dopo l’aggiunta di acido retinoico 10-9 M (De Luca, 1991). Fortunatamente, la capacità dell’acido retinoico di revertire il fenotipo neoplastico di differenti linee cellulari è spesso riproducibile anche in vivo: RA è infatti efficace nel prevenire la comparsa di forme tumorali chimicamente indotte in animali in diversi tessuti (pelle, intestino, vescica, mammella), e test clinici sono in atto per valutare anche nell’uomo la presenza di analoghi effetti (Lippman et al., 1995). Già oggi l’acido retinoico viene impiegato con successo nel trattamento di alcuni tumori cutanei e del tratto aero-digestivo, ed estremamente promettenti sono i risultati nel trattamento di neoplasie del tessuto ematopoietico, quali ad esempio la leucemia acuta promielocitica (Smith et al., 1992; Cornic, et al., 1994a).La morte programmata delle cellule (apoptosi) può essere considerata un particolare fenomeno differenziativo, che sembra essere mediata, in tessuti adulti ed embrionali, da endonucleasi Ca++/ Mg++ dipendenti. L’acido retinoico induce questo processo in tessuto mesenchimale di embrioni di topo, in granulociti neutrofili cresciuti in vitro, o in alcune linee cellulari tumorali, tra le quali le cellule leucemiche HL-60 (Martin et al., 1990.Un’altra importante azione biologica esercitata dall’acido retinoico è quella del controllo dello sviluppo embrionale nei vertebrati: RA è un potente morfogeno, indispensabile per il corretto sviluppo dello scheletro, degli arti, e degli organi interni (Gudas, 1994), ma a dosi farmacologiche è anche un potente agente teratogeno, che provoca gravi malformazioni ossee, del tessuto epidermico, e nell’organogenesi, in quanto attraversa facilmente la barriera placentare, e tende ad accumularsi nei tessuti embrionali (Tzimas et al., 1994).L’acido retinoico è in grado di sostituire il retinolo in tutte le sue funzioni, ad eccezione delle funzioni visive e, in alcune specie, di quelle riproduttive, evidenziando sempre una maggiore potenza ed efficacia. Infatti, quando RA è somministrato nella dieta come unico retinoide, gli animali crescono normalmente, ma sono ciechi (Dowling e Wald, 1960), e si ha assenza di spermatogenesi, o necrosi placentare (Thompson et al., 1969). Si può affermare quindi con buona approssimazione che la quasi totalità degli effetti biologici attribuiti al retinolo siano in realtà conseguenti alla sua trasformazione metabolica nella molecola biologicamente attiva, l’acido retinoico, e determinati quindi dall’azione di attivazione genica di cui quest’ultimo composto è capace.
5

Introduzione
2.2 MECCANISMO D'AZIONE NUCLEARE DELL'ACIDO RETINOICO
L’isolamento, nel 1987, da parte di Giguere et al., e Petkovich et al. dei primi recettori nucleari (retinoic acid receptors, RARcapaci di attivare la trascrizione in seguito al legame con l’isomero all-trans dell’acido retinoico (all-trans RA) (Kdiss: 1-5 nM), ha definitivamente confermato l’ipotesi che tale molecola agisse controllando in modo specifico il processo di trascrizione in modo del tutto analogo a quello degli ormoni steroidei, ed ha aperto la via verso l’individuazione dei geni attivati da RA e dei meccanismi molecolari con cui esso esercita i suoi molteplici effetti (Giguere et al., 1987; Petkovich et al., 1987). Fino ad oggi, tre diversisottotipi di RAR (e tutti appartenenti alla superfamiglia dei recettori nucleari degli ormoni steroidei, sono stati identificati in mammiferi, uccelli e anfibi, l’espressione dei quali risulta essere strettamente tempo e tessuto dipendente (Tabella 1) (Kastner e Chambon, 1994).
RecettoreCromosoma
(uomo)
Pattern di espressione
Embrione Adulto
RAR 17 Ubiquitario Ubiquitario
RAR 3 Specifico
(Muscolo, Prostata)Spazio e tempo dipendente
RAR 12 Specifico
(Pelle, Intestino)Spazio e tempo dipendente
RXR 9 Specifico
(Pelle, Fegato, Reni)
Specifico
(Intestino, Pelle, Fegato)
RXR 6 Ubiquitario Ubiquitario
RXRg 1 Specifico
(Muscoli, Cuore)
Specifico
(Cervello, Ipofisi)
Tabella 1: Localizzazione cromosomica e pattern di espressione di RAR e RXR in differenti tessuti umani
Le RAR (P.M.= 50000) possiedono 6 distinti domini strutturali (A-F) funzionalmente importanti per il legame con RA, con il DNA, per la dimerizzazione con altri recettori o per l’interazione con l’apparato di trascrizione. La conservazione della sequenza amminoacidica tra le tre isoforme è notevole - oltre il 75% di omologia - in particolare nella regione che lega l’acido retinoico e in quella che interagisce con il DNA (Kastner e Chambon 1994).
6

Introduzione
Un’altra importante scoperta è stata quella dell’isolamento, nel 1990, di una nuova classe di proteine appartenenti alla superfamiglia dei recettori degli ormoni steroidei, i retinoid X receptors (RXR, sottotipi e ), che presentano tuttavia una bassa omologia di sequenza amminoacidica nei confronti delle RAR (Mangelsdorf, et al., 1990). Le tre distinte isoforme di RXR (P.M.: 50000) possiedono, come le RAR sei domini funzionalmente distinti, ed hanno patterns di espressione temporalmente e spazialmente dipendenti (Mangelsdorf et al., 1994).Il ligando naturale di RXR è stato identificato solo recentemente, nel 1992: è l’acido 9-cis retinoico (9-cis RA), un composto del quale fino a quel momento non era noto il ruolo fisiologico, e la cui origine rimane tuttora ignota (Figura 2).
Figura 2: Formule di struttura di acido 9-cis e all-trans retinoico
Questo isomero dell’acido retinoico (che nella forma all-trans lega debolmente le RXR), è un potente attivatore trascrizionale delle tre isoforme di RXR, che lega con elevata affinità (Kdiss: circa 10 nM) (Heyman et al., 1992; Levin, et al., 1992).
L’identificazione di questa nuova classe di recettori nucleari leganti un particolare isomero dell’acido retinoico ha rappresentato il primo passo verso la comprensione delle modalità con cui una molecola così semplice possa mediare una tale varietà di effetti biologici.I recettori nucleari degli ormoni steroidei si legano al DNA sotto forma di eterodimeri, i quali, in presenza dei rispettivi ligandi, subiscono modificazioni conformazionali che portano all’attivazione o all’inibizione della trascrizione dei geni bersaglio. Il legame dei recettori con uno o entrambi i ligandi determina una risposta quantitativamente differente.
7

Introduzione
Numerosi esperimenti effettuati in vitro e in vivo hanno dimostrato che le diverse forme di RXR hanno un ruolo cruciale nel complesso sistema di regolazione dell’espressione genica, in quanto essi costituiscono i partners necessari per la dimerizzazione e il legame con il DNA non solo delle RAR, ma anche di tutti i recettori nucleari appartenenti alla sottofamiglia di tipo II. Tra questi vi è il recettore dell’ormone tiroideo (TR), della vitamina D (VDR), il peroxisome proliferator-activated receptor (PPAR), che lega acidi grassi, oltre ad una serie di recettori nucleari dei quali non è stato ancora identificato il ligando (Leid et al., 1993).E’ quindi probabile che il 9-cis RA, attraverso il legame con RXR, possa avere un ruolo fondamentale nel modulare la trascrizione non solo dei geni sensibili ad all-trans RA, ma anche di tutti quelli regolati dai ligandi e dai recettori sopracitati: questa possibile importante funzione rende ancora più importante individuare l’origine biologica di tale molecola.Il legame dei recettori con il DNA avviene grazie al riconoscimento di specifiche sequenze localizzate a monte del gene bersaglio denominate, nel caso delle due classi di recettori nucleari dell’acido retinoico, RAR e RXR response elements (RAREs e RXREs). Queste sequenze sono formate dalla ripetizione in forma diretta, o come palindrome, di una sequenza esamerica AGGTCA, separata da 1 a 5 nucleotidi (Rastinejad et al., 1995; Kurokawa et al., 1994). La presenza a monte di un gene bersaglio di questi differenti siti di consenso seleziona quindi quale tra i possibili eterodimeri si possa legare, quale sia la disposizione spaziale relativa (polarità) dei due singoli recettori, e, di conseguenza, il tipo di modulazione trascrizionale (positiva o negativa) indotta sul gene bersaglio in presenza dei ligando.Ad esempio, la presenza di una sequenza di consenso di tipo DR +5 (in cui cioè si ha ripetizione diretta della sequenza esamerica separata da 5 nucleotidi) a monte di un gene bersaglio, determina il legame del dimero RAR/RXR, e l’attivazione della trascrizione in seguito al legame con 9-cis e/o all-trans RA, mentre una sequenza di tipo DR +1 lega lo stesso eterodimero, ma con polarità opposta, determinando inibizione della trascrizione in presenza del/i ligando/i (Kurokawa et al., 1995; Figura 3).
AGGTCA AGGTCA5 Gene X
RXR RAR
9-cis RA
Attivazione dellaespressione genica
All-trans RA
DNARARE AGGTCA AGGTCA1 Gene Y
RAR RXR
Inibizione dellaespressione genica
All-trans RA 9-cis RA
RARE DNA
Figura 3: Schema del meccanismo d’azione dei recettori nucleari dell’acido retinoico
8

Introduzione
Il meccanismo di modulazione della trascrizione genica da RA è comunque estremamente complesso, ed è stato chiarito solo parzialmente: in particolare sono ancora oscuri i meccanismi di interazione dei recettori nucleari con i componenti dell’apparato di trascrizione; a questo proposito è interessante rilevare che sono stati identificati di recente alcuni componenti proteici (N-CoR, p140 e p160) che, dissociandosi dalle RAR o associandosi ad esse in seguito al legame di all-trans RA, fungono da co-attivatori o co-repressori della trascrizone (Horlein et al., 1995; Kurokawa et al., 1995).A causa della notevole instabilità di queste proteine, la struttura di RAR e RXR è stata fino ad ora risolta solo parzialmente. Nel caso di RXRad esempio, è stata determinata la struttura del dominio responsabile del legame con 9-cis RA e della dimerizzazione, e di quello che lega il DNA (Lee et al., 1993; Rastinejad et al., 1995). L’identificazione della struttura completa di RXR e RAR permetterà un decisivo passo avanti nella comprensione dei meccanismi di attivazione trascrizionale e delle modalità di interazione intermolecolari di queste proteine. I geni sino ad oggi identificati, che contengono “a monte” una sequenza RARE o RXRE sono circa un centinaio: la maggior parte di questi codificano per proteine coinvolte nel metabolismo del retinolo (e dell’acido retinoico), o sono geni importanti per i processi di proliferazione o differenziamento (Stunnenberg, 1993; Smith et al., 1992). Tra i primi, troviamo alcuni tra gli stessi recettori nucleari (RAR , le proteine intracellulari leganti il retinolo o l’acido retinoico (Cellular retinol-binding protein, CRBP tipo I e II; Cellular retinoic acid-binding protein, CRABP tipo II) e deidrogenasi forse coinvolte nel processo di sintesi di RA (alcool deidrogenasi di classe 3, ADH-3) (Sukov et al., 1990; Smith et al., 1991; Giguere et al., 1990; Duester et al., 1991). Tra i geni importanti per il mantenimento della crescita e il differenziamento, contenenti “a monte” sequenze di consenso RARE o RXRE, vi sono gli homeobox genes, un gruppo di fattori di trascrizone essenziali, nello sviluppo embrionale, per la formazione degli arti (Scott e Goldsmith, 1993), l’ormone della crescita, indotto nell’ipofisi in modo sinergico da RA e dall’ormone tiroideo (Bedo et al., 1989), il gene codificante per la laminina B1, importante per l’induzione del differenziamento nelle cellule di carcinoma embrionale F9 (Vasios et al., 1989), oltre alle proteine osteocalcina e osteopontina, coinvolte nella regolazione del ciclo e della divisione cellulare (Schule et al., 1990).
2.3 I RETINOIDI COME FARMACI
L’acido retinoico ed i suoi derivati di sintesi sono da alcuni anni diventati composti di elevato interesse farmacologico grazie alla grande efficacia dimostrata nel trattamento di numerose patologie, principalmente in dermatologia e in terapia antineoplastica.L’acido all-trans retinoico, somministrato per via orale, è stato inizialmente impiegato con successo nel trattamento di forme gravi di acne (Es. acne vulgaris, acne
9

Introduzione
nodulocistica), grazie alla sua capacità di ridurre la dimensione e la secrezione delle ghiandole sebacee (Kligman et al., 1969). Si è inoltre dimostrato efficace, mediante somministrazione per via topica, nel ridurre l’iperproliferazione e nel regolarizzare il differenziamento della cute in varie forme di psoriasi, tra le quali l’artropatia psoriasica; notevole è la sua attività nel favorire il riparo della pelle fotodanneggiata da UV, e, in cosmesi, quale principio attivo di alcune creme antirughe (Weiss et al., 1988; Kligman, 1986).Sebbene siano stati ottenuti risultati notevoli nel trattamento di varie dermatosi, non meno importanti sono le applicazioni dell’acido retinoico nella prevenzione e il trattamento di varie forme neoplastiche.L’acido retinoico è capace di prevenire la ricomparsa di tumori secondari del tratto digestivo e respiratorio (numerosi test clinici sono in atto per valutare l’efficacia anche in altri tipi di neoplasie) (Vahlquist e Rollman, 1987). L’acido all-trans retinoico, somministrato oralmente, si dimostra efficace nel trattamento di alcuni tumori della pelle (Es. Xeroderma pigmentoso, carcinoma delle cellule basali, melanoma) (Epstein, 1986), e nel revertire lesioni neoplastiche del tratto oro-faringeo (leucoplachia orale, papillomi laringei) (Lippmann et al., 1987), ma è soprattutto nel trattamento della leucemia acuta promielocitica (APL) che sono stati ottenuti i risultati più promettenti (Breitman et al., 1980). Questa forma di leucemia corrisponde a circa il 10% dei casi di leucemia mieloide, ed è provocata dalla presenza del sangue di promielociti immaturi, a causa di una traslocazione cromosomica, nei cromosomi 15 e 17, tra il gene codificante per una proteina detta PML (un fattore di trascrizione che contribuisce tra l’altro alla formazione di un complesso multiproteico nucleare) (Borden et al., 1995), e il gene codificante per RAREarlye Dimitrovsky, 5. La proteina di fusione PML/RAR così formata, ha presumibilmente un ruolo determinante nella genesi del processo neoplastico, agendo da effettore negativo dominante sul differenziamento dei promielociti. La terapia mediante somministrazione orale di all-trans RA è associata a completa remissione (CR) in oltre il 90% dei pazienti affetti da APL (dopo circa 1-2 mesi di trattamento), inducendo il differenziamento dei promielociti a granulociti maturi attraverso un meccanismo tuttora sconosciuto (Grignani et al., 1994; Muindi, 1994). Il periodo di CR in seguito al trattamento con solo all-trans RA è di durata variabile (4-18 mesi), significativamente maggiore (in alcuni casi si ha guarigione definitiva) se la somministrazione di all-trans RA è associata con chemioterapia convenzionale (Chomienne, 1994). E’ stato dimostrato che la ricomparsa dei sintomi è causata da un’insufficiente concentrazione plasmatica di all-trans RA, il quale induce il proprio catabolismo e la sintesi della proteina legante CRABP nei tessuti: in questo caso la somministrazione RA, anche a dosi elevate, non si dimostra efficace per il trattamento della malattia. (Cornic, 1994b). Le principali linee di ricerca sono rivolte alla comprensione del ruolo giocato dalla proteina PML/RAR nella patogenesi di APL, e del meccanismo molecolare d’azione con
10

Introduzione
cui all-trans RA reverte il fenotipo neoplastico. La possibilità di controllare la biosintesi di RA nei tessuti, l’impiego di farmaci che blocchino il suo catabolismo (Schwartz et al., 1995), o la scoperta di nuovi retinoidi naturali e di sintesi più stabili e attivi potranno costituire efficaci strumenti per ottenere un definitivo successo nel trattamento della leucemia acuta promielocitica.Uno degli ostacoli maggiori all’impiego terapeutico di sostanze ad elevata attività biologica e ad ampio spettro d’azione quali l’acido retinoico, è costituito dal basso indice terapeutico. Alle dosi comunemente impiegate, frequenti sono gli effetti collaterali: si può manifestare sindrome da ipervitaminosi A, con sintomi a volte gravi a livello del sistema nervoso centrale, dell’apparato digerente e osteoarticolare, così come frequenti sono le infiammazioni ed alterazioni dei tessuti cutanei. Notevoli sono inoltre le limitazioni imposte dalla potente attività teratogena ed embriotossica dell’acido retinoico (Tzimas et al., 1994; Smith et al.,1992). Quando somministrato per via orale, l’acido retinoico ha una bassa biodisponibilità (30%), e un rapido t1/2 (30 min.) (Muindi, 1994). La farmacocinetica di RA è estremamente complessa poichè, essendo una molecola naturale, forma complessi con le proteine plasmatiche, e, nel caso di somministrazioni prolungate, attiva il proprio catabolismo rendendo estremamente difficile prevedere le curve di concentrazione plasmatica.Per queste ragioni sono intensamente ricercati analoghi naturali o di sintesi di all-trans RA, che possiedano la medesima attività biologica, minore tossicità, e migliori parametri farmacocinetici. L’acido 9-cis retinoico, retinoide naturale scoperto recentemente, capace di attivare selettivamente RXR, è ancora scarsamente impiegato in terapia, ma i suoi effetti sono oggetto di intenso studio. L’attività biologica di 9-cis RA (CR è osservata nel trattamento di APL), così come i suoi parametri farmacocinetici, non sono distinguibili da quelli determinati quando è somministrato all-trans RA, presumibilmente perchè, alle dosi impiegate, avviene nel plasma una consistente isomerizzazione di tale composto nella forma isomerica all-trans (Doran, 1994; Miller et al., 1995). Nella Figura 4 sono riportate le formule di struttura di alcuni tra i principali retinoidi di sintesi dotati di interessanti attività biologiche, disponibili commercialmente o attualmente in fase di test clinici. Sono comunque tutte sostanze, sebbene spesso estremamente efficaci, la cui somministrazione crea problemi di tossicità e teratogenicità, in modo più o meno rilevante a secondo dei casi.Tra questi, il 13-cis acido retinoico (isotretinoina), è oggi il retinoide preferito per la somministrazione sistemica nel trattamento di acne grave, in quanto sono stati evidenziati minori effetti collaterali rispetto all’impiego di all-trans RA (Boyd, 1989); l’isotretinoina possiede inoltre attività chemiopreventiva, in quanto, se somministrata a pazienti con neoplasie della cavità orale, inibisce significativamente la formazione di tumori secondari (Lippman et al., 1995).
11

Introduzione
L’etretinate e il suo metabolita acitretina sono retinoidi impiegati oggi come prima scelta per il trattamento di tutte le forme di psoriasi e ittiosi, disordini della iperproliferazione e della cheratinizzazione della cute. Essendo sostanze altamente lipofile, entrambe si accumulano a lungo nei tessuti, mantenendo elevato il rischio di teratogenesi anche per anni (Fritsch, 1992).Il composto 4-idrossifenilretinamide (4-HPR) è uno dei retinoidi più promettenti tra quelli non ancora disponibili commercialmente, in quanto si è dimostrato efficace nella prevenzione di svariati tumori indotti chimicamente su animali; sono in fase avanzata di sperimentazione sull’uomo test clinici (fase III) per valutare la sua efficacia nel prevenire la ricomparsa di tumori al seno (Formelli et al.,1993).
12

Introduzione
Figura 4: Formule di struttura di differenti retinoidi di impiego farmacologico
Poichè in alcuni casi è stata osservata una buona correlazione tra l’attività farmacologica dei retinoidi somministrati e la loro affinità per la CRBPI (Hill e Grubbs, 1992), abbiamo verificato le proprietà di legame del retinil metil etere, composto di sintesi che è risultato efficace in differenti test clinici nella prevenzione di tumori al seno (Shealy, 1989). In effetti questo retinoide, strutturalmente identico al retinolo, se si eccettua il gruppo metilico terminale, lega con buona affinità la CRBPI, con una Kdiss apparente di 2 nM,
che è circa 20 volte maggiore di quella del ligando endogeno, l’all-trans retinolo (Malpeli et al., 1996).Sono state recentemente identificate alcune molecole di sintesi, agonisti ed antagonisti selettivi, capaci di modulare, in vitro, la risposta trascrizionale nelle due distinte classi di recettori nucleari per l’acido retinoico (Figura 5) (Kiedel et al., 1994; Fanjui et al., 1994). Sarà estremamente interessante valutare gli effetti biologici in vivo di tali composti, che potrebbero essere i capostipiti di una nuova classe di retinoidi dotati di una maggiore selettività e specificità d’azione, oltre che di una maggiore stabilità.
Figura 5: Agonisti selettivi di RAR e RXR
2.4 ASSUNZIONE E TRASPORTO DI RETINOLO
2.41 Assorbimento intestinale di retinoidi e metabolismo epatico
La dieta costituisce l’unica fonte di provenienza per l’organismo dei retinoidi, che sono assunti come carotenoidi di origine vegetale (per lo più sotto forma di all-trans -carotene), o come retinil esteri o retinolo di origine animale.I retinoidi sono assorbiti prevalentemente sotto forma di retinolo, in quanto i retinil esteri vengono idrolizzati nel lume del piccolo intestino da parte di idrolasi pancreatiche non specifiche, o da di retinil estere idrolasi presenti sui villi degli enterociti intestinali
13

Introduzione
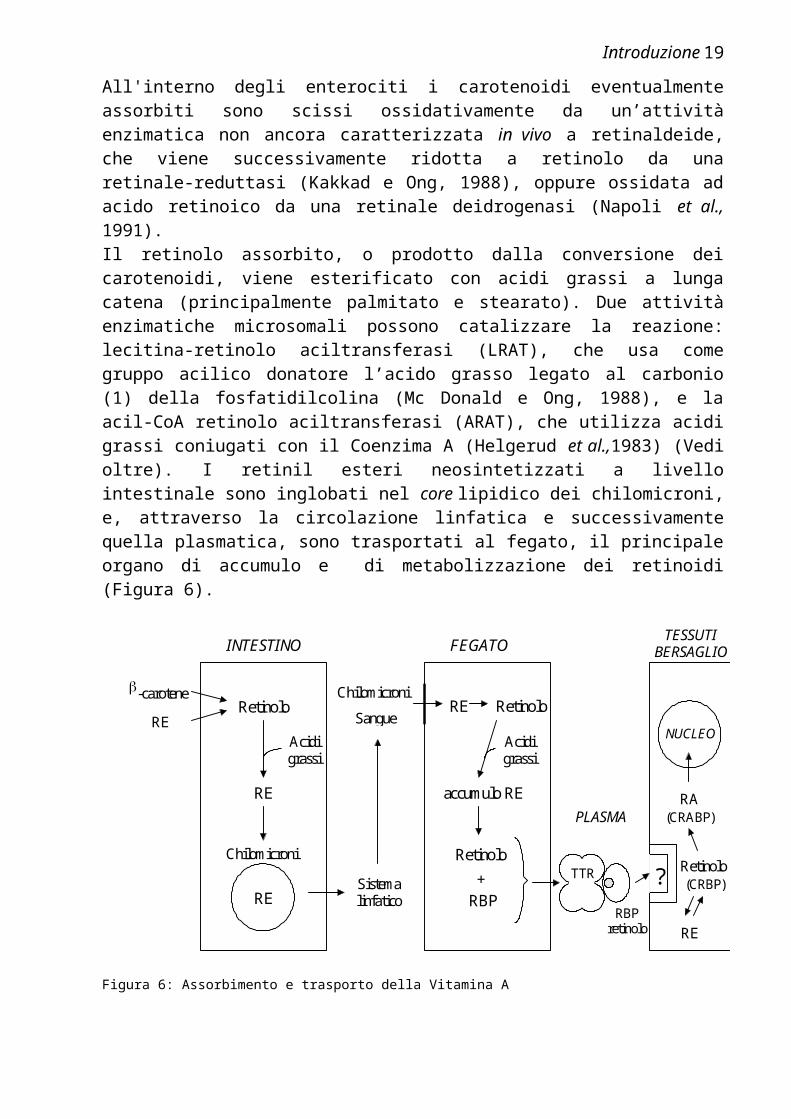
(Rigtrup e Ong, 1992). L’assorbimento dell’acido retinoico a livello intestinale è possibile, ma quantitativamente trascurabile, poichè RA è presente solo in tracce nei cibi (Fidge et al., 1968).Abbiamo potuto verificare sperimentalmente l’importanza dell’assorbimento intestinale per il mantenimento dei normali livelli di Vitamina A nell’organismo analizzando i livelli plasmatici di retinolo in un individuo obeso che, per eliminare questo problema, si era sottoposto ad un intervento di rimozione di una parte consistente dell’intestino tenue, ma, ad alcuni mesi dall’intervento, mostrava sintomi evidenti di ipovitaminosi A (cecità notturna), nonostante avesse mantenuto una dieta normale ed avesse ancora problemi di sovrappeso. L’analisi effettuata ha confermato l’esaurimento pressochè totale delle riserve di Vitamina A dell’organismo, evidenziando la presenza di una concentrazione plasmatica di ROH che era solo il 4% di quella di un individuo normale.All'interno degli enterociti i carotenoidi eventualmente assorbiti sono scissi ossidativamente da un’attività enzimatica non ancora caratterizzata in vivo a retinaldeide, che viene successivamente ridotta a retinolo da una retinale-reduttasi (Kakkad e Ong, 1988), oppure ossidata ad acido retinoico da una retinale deidrogenasi (Napoli et al., 1991).Il retinolo assorbito, o prodotto dalla conversione dei carotenoidi, viene esterificato con acidi grassi a lunga catena (principalmente palmitato e stearato). Due attività enzimatiche microsomali possono catalizzare la reazione: lecitina-retinolo aciltransferasi (LRAT), che usa come gruppo acilico donatore l’acido grasso legato al carbonio (1) della fosfatidilcolina (Mc Donald e Ong, 1988), e la acil-CoA retinolo aciltransferasi (ARAT), che utilizza acidi grassi coniugati con il Coenzima A (Helgerud et al.,1983) (Vedi oltre). I retinil esteri neosintetizzati a livello intestinale sono inglobati nel core lipidico dei chilomicroni, e, attraverso la circolazione linfatica e successivamente quella plasmatica, sono trasportati al fegato, il principale organo di accumulo e di metabolizzazione dei retinoidi (Figura 6).
14

Introduzione
RBPretinolo
?
-carotene
RERetinolo
INTESTINO FEGATOTESSUTI
BERSAGLIO
RE
RESistemalinfatico
Sangue
Chilomicroni
accumulo RE
Retinolo +
RBP
PLASMA
Acidigrassi
Retinolo
RE
RA
NUCLEO
(CRABP)
(CRBP)TTR
Acidigrassi
Chilomicroni
RE Retinolo
Figura 6: Assorbimento e trasporto della Vitamina A
Una minima porzione dei retinoidi assunti con la dieta è presente nel plasma come acido retinoico legato ad albumina (non è stato ancora identificato alcun trasportatore plasmatico specifico), ad una concentrazione massima (4-14 nM nell’uomo) pari all’1% di quella del retinolo. Si ritiene che l’acido retinoico del plasma non contribuisca in modo significativamente rilevante al pool di RA intratissutale, principalmente formato dall’ossidazione in situ del retinolo (Blaner et al.,1994a).Il processo di accumulo e mobilizzazione dei retinoidi nel fegato è estremamente complesso, e non ancora del tutto chiarito. I retinil esteri trasportati dai chilomicroni sono accumulati nelle cellule stellate epatiche in seguito ad un ciclo di idrolisi e successiva esterificazione (Blohmoff et al., 1984). Essi costituiscono la riserva principale di retinoidi dell’organismo, sebbene altri tessuti (tessuto adiposo, intestino, reni) contengano retinil esteri in quantità significativa (Blaner et al., 1994b).
2.42 Trasporto plasmatico di retinolo: la RBP
Il retinolo, mobilizzato in seguito all’idrolisi dei retinil esteri, viene secreto dal fegato in forma legata alla Retinol Binding Protein (RBP), la proteina che ha la funzione di trasportarlo nel plasma e di rilasciarlo ai tessuti bersaglio.La RBP (P.M.= 21000) interagisce nel plasma con la transtiretina (TTR, o prealbumina, P.M.= 55000) a formare un complesso che, incrementando le dimensioni molecolari relativamente modeste della RBP, sembra prevenirne la filtrazione glomerulare (Berni et al., 1993; Soprano e Blaner, 1994). La struttura molecolare del complesso costituito da retinolo, RBP e TTR (presenti in rapporto 2:2:1) è stata recentemente risolta (Monaco et al., 1995; Figura 7).
15

Introduzione
Figura 7: Modello di struttura del complesso esamerico (oloRBP)2-TTR (da Monaco et al., 1995). Al centro il tetramero di TTR, all’esterno la RBP con il retinolo legato.
La concentrazione della oloRBP nel plasma è strettamente regolata, ed è mantenuta costante (2-3 M), principalmente mediante il controllo della sintesi di RBP nelle cellule parenchimali (Soprano e Blaner, 1994).La RBP umana è costituita da un -barrel formato da due foglietti, con otto catene -antiparallele e una corta elica, che permette il legame con una sola molecola di retinolo, il quale si inserisce profondamente nella cavità di legame con l’anello cicloesenico rivolto verso l’interno e il gruppo ossidrile esposto al solvente (Cowan et al., 1990; Figura 7). Sebbene il ligando endogeno sia all-trans retinolo, la RBP lega in vitro con simile affinità (Kdiss: circa 10-7 M), gli isomeri 13- e 11-cis del retinolo, la
retinaldeide, l’acido retinoico, ed altri composti con differenti gruppi funzionali, mentre non ha affinità per i retinoidi modificati nell’anello cicloesenico (Cogan et al., 1976; Berni et al., 1993; Zanotti et al., 1993).
2.43 Il rilascio del retinolo ai tessuti
Il rilascio del retinolo ai tessuti determina la dissociazione del complesso RBP-TTR, e la filtrazione glomerulare della RBP (Valquist et al., 1973). Sono state proposte due ipotesi riguardo alle modalità del rilascio e della assunzione intracellulare del retinolo, processi che non sono stati ancora completamente chiariti. Bok e Heller (Bok e Heller, 1976) hanno dimostrato l’esistenza di un complesso oloRBP-recettore sulla membrana cellulare di epitelio pigmentato di retina. Recentemente è stato purificato un recettore di 63 KDa da frazioni microsomali di epitelio retinico che è in grado di legare apo e olo RBP, ma nessuna evidenza funzionale che sia effettivamente
16

Introduzione
coinvolto nel processo di uptake di retinolo, piuttosto che in altri (Es.: degradazione della RBP) è stata finora fornita (Bavik et al., 1991). Sebbene l’ipotesi del recettore sia attraente, un’altra proposta è che il retinolo diffonda dalla RBP plasmatica attraverso la membrana cellulare delle cellule bersaglio, e si leghi direttamente alla apoCRBP all’interno dei tessuti, mediante un processo la cui velocità è determinata dalla concentrazione di accettore in forma apo (Noy e Blaner, 1991). Questa ipotesi è avvalorata dal fatto che, incubando oloRBP in presenza di liposomi o membrane cellulari, è stata osservata dissociazione spontanea del retinolo. Questo fenomeno è causato forse dalla micro-denaturazione reversibile della proteina indotta dal pH acido presente in prossimità delle membrane (Bychkova et al., 1992).
2.5 METABOLISMO INTRACELLULARE DEL RETINOLO
2.51 I trasportatori intracellulari dei retinoidi
A causa della loro scarsa solubilità, instabilità chimica, e delle loro proprietà membranolitiche, tutti i retinoidi, ad eccezione dei retinil esteri, sono sempre associati a trasportatori proteici durante il trasporto extracellulare o intracellulare. Tali proteine non hanno solo una funzione “protettiva” o di “immagazzinamento” dei retinoidi, ma sono altresì fondamentali nel dirigere il metabolismo dei propri ligandi, promuovendone l’accesso ad enzimi specifici mediato, molto probabilmente, da riconoscimento selettivo proteina-proteina (Tabella 2).
Binding Protein Retinoide legato Localizzazione
RBP All-trans retinolo Fegato, tessuti extraepatici
CRBPI All-trans retinolo Ubiquitaria; fegato, reni, intestino, occhi, milza
CRBPII All-trans retinolo Piccolo intestino
CRABPI Acido all-trans retinoico
Ubiquitaria; vescicole seminali, vas deferens, pelle, occhi
CRABPII Acido all-trans retinoico
Pelle
Tabella 2: Ligando endogeno e localizzazione tissutale delle principali retinoid-binding proteins
Mentre la RBP è l’unico carrier extracellulare del retinolo, quattro sono i principali dei trasportatori intracellulari dei retinoidi: CRBPI e CRBPII, proteine leganti il retinolo, e CRABPI e II, proteine leganti l'acido retinoico. Tutte fanno parte della famiglia genica
17

Introduzione
Figura 8: Stereogramma della sovrapposizione del modello di struttura a carboni tra CRBPI, CRBPII (grigio) e fatty acid-binding protein di insetto (nero)
delle CRBP/FABP, che comprende 11 proteine omologhe, trasportatori di piccole molecole idrofobiche, incluse alcune fatty acid binding proteins, e la proteina mielinica P2. Le proteine della famiglia presentano notevoli somiglianze di sequenza amminoacidica e strutturali. Esse sono costituite in media 135 amminoacidi, e possiedono una identità a livello di struttura primaria variabile dal 40% al 78% (Banaszak et al., 1994).Tali analogie nella sequenza amminoacidica trovano riscontro in una elevata somiglianza strutturale. Infatti, anche quando l’identità di sequenza primaria è bassa (Es. 10% tra CRBPI e FABP di insetto) si osserva una notevole sovrapponibilità conformazionale delle strutture secondarie (Figura 8; Newcomer, 1995).La CRBP e la CRABP (tipo I e II) sono costituite da 10 catene antiparallele, disposte in modo da formare un “-barrel”. Nella cavità di legame, il ligando si presenta con il gruppo funzionale rivolto verso l’interno, in modo opposto a quanto avviene nella RBP. Due corte -eliche chiudono l’apertura del -barrel rendendo l’anello cicloesenico del ligando pressochè inaccessibile al solvente (Figura 9, 10; Newcomer, 1995).Nonostante la notevole somiglianza strutturale e di sequenza, le retinoid-binding proteins intracellulari differiscono notevolmente per affinità e specificità di legame, oltre che per il ruolo fisiologico completamente differente. I due tipi di proteine (I e II) hanno inoltre distinta localizzazione tissutale, essendo la CRBP I e la CRABP I ubiquitarie, e particolarmente abbondanti in fegato, epiteli, testicolo, rene, polmoni, mentre la presenza delle forme II è ristretta agli enterociti (CRBPII), ed alle cellule epidermiche (CRABPII) (Tabella 2).
18
Introduzione
La CRBPI (P.M.= 14700; Figura 9, 10) lega con elevata affinità il ligando endogeno all-trans retinolo (Kdiss @ 0.1 nM), e, con un’affinità di due ordini di grandezza minore, la
all-trans retinaldeide e il 13-cis retinolo (Li et al., 1991). I dati riportati in letteratura indicano l’assoluta mancanza di affinità della CRBPI per qualsiasi isomero dell’acido retinoico, così come per il 9-cis retinolo (Mc Donald e Ong, 1987). Nella cavità di legame gioca un ruolo essenziale nel determinare la specificità di legame la catena laterale dell’amminoacido glutamina 108, che forma un legame idrogeno con il gruppo ossidrile del retinolo (Cowan et al., 1993). Nelle retinoid-binding proteins intracellulari, contrariamente a quanto avviene nella RBP, modifiche nel gruppo funzionale terminale possono determinare anche l’abolizone del legame, mentre l’affinità non viene alterata in modo drastico per retinoidi modificati sull’anello cicloesenico (Ong e Chytil, 1988). Un aspetto non ancora chiarito è quello del meccanismo di rilascio e del legame del retinolo dal trasportatore cellulare. L'inaccessibilità al solvente del ligando, unitamente all'identità strutturale fra la CRBP in forma apo e olo, rendono necessario ipotizzare l'esistenza di una importante transizione strutturale della proteina: l’ipotesi più plausibile è che il -barrel si “apra” in corrispondenza delle uniche due catene antiparallele del foglietto (D e E) (Figura 9, b) che non formano tra loro legami ad idrogeno, e che tra l’altro si trovano in prossimità del ligando (Banaszak et al., 1994).
Figura 9: Struttura della oloCRBPI rappresentata secondo il modello a “spazio pieno” (a), o visualizzando i carboni bIn (b) la freccia indica la posizione delle due catene “D” ed “E”, attraverso le quali potrebbe avvenire il rilascio del retinolo.
19

Introduzione
Il complesso retinolo-CRBP costituisce la forma prevalente nella quale è presente il retinolo all’interno delle cellule, che viene così preservato da reazioni metaboliche aspecifiche, e reso disponibile come substrato per specifici enzimi nei processi di esterificazione o di ossidazione ad acido retinoico.La CRBPI in forma apo è presente nel citoplasma delle cellule di vari tessuti, in quanto il rapporto molare stimato retinolo:CRBPI è minore di 1 (Harrison et al., 1987). Tale rapporto, variabile in modo dinamico a seconda delle condizioni nutrizionali dell’animale, potrebbe costituire un importante fattore per la regolazione dei principali processi metabolici di conversione del retinolo. Tra l’altro, la presenza di apoCRBP all’interno delle cellule contribuisce in modo determinante a minimizzare la concentrazione di retinolo libero in equilibrio.La CRBPII (P.M.= 15600) (Winter et al., 1993) possiede un’identità di sequenza nei confronti della CRBPI del 56%, e una pressochè perfetta sovrapponibilità strutturale (Figura 8), ma la affinità nei confronti di all-trans retinaldeide e all-trans retinolo è all’incirca la stessa, e la Kdiss per quest’ultimo è maggiore di due ordini di grandezza
rispetto a quella della CRBPI (Li et al., 1991). La cavità di legame della CRBPII ha caratteristiche simili a quella della CRBPI. Nonostante siano stati effettuati diversi esperimenti di mutagenesi sito-specifica, non è stato ancora chiarito quali amminoacidi siano responsabili della differente specificità di legame tra la CRBPI e la CRBPII (Jamison et al., 1995). Forse la mancata discriminazione nel legame con ROH e RALD di quest’ultima è dovuta a una differente disposizione spaziale del ligando nella formazione del legame idrogeno con la glutamina in posizione 108 (Newcomer, 1995).La CRBPII, per la sua specifica localizzazione, è coinvolta nei processi metabolici correlati con l’assorbimento intestinale dei retinoidi. Il complesso CRBPII-retinaldeide, formatosi in seguito alla scissione ossidativa del -carotene, è il substrato di una reduttasimicrosomale presente nella mucosa intestinale che catalizza la formazione di retinolo (Kakkad e Ong, 1988). Quest’ultimo, legato alla CRBPII, viene successivamente esterificato dall’attività LRAT, sempre nel tessuto intestinale (Herr e Ong, 1992)La CRABPI (P.M.= 14700; Figura 10) è la più abbondante tra le proteine leganti l'acido retinoico. Ha una costante di dissociazione di dell’ordine di 10-9 M per all-trans RA, ed è in grado di legare 9-cis RA e 13-cis RA, sebbene con affinità circa 50 e 150 volte minore, rispettivamente.La CRABPI non lega il retinolo o la retinaldeide (Fiorella et al., 1993). Il legame dell’acido retinoico avviene in posizione leggermente più esterna rispetto a quanto osservato per la CRBP, e riveste un ruolo essenziale per il legame la presenza delle catene laterali di arginina in posizione 111 e 131, che formano legami idrogeno con il gruppo carbossilico dell’acido retinoico (Kleywegt et al., 1994; Figura 10).Esperimenti di mutagenesi sito-specifica effettuati su CRBPI e CRABPI, che consistevano nello scambio degli amminoacidi ritenuti indispensabili per il legame (Gln 108 e Arg 111 e 131, rispettivamente), hanno mostrato che le proteine mutate hanno una
20

Introduzione
ridotta o quasi nulla affinità per il ligando fisiologico, e non sono in grado di legare nuovi retinoidi. Questo dato suggerisce che, oltre ai legami appena indicati, altre interazioni tra ligando e amminoacidi della cavità di legame sono importanti nel conferire la specificità di legame alla proteina (Stump et al., 1991; Zhuang et al., 1992). La CRABPI e II hanno il 78% di identità di sequenza primaria, mentre tra CRABPI e CRBPI l’identità è del 40%, ma tra le tre proteine vi è una notevole sovrapponibilità strutturale (Banaszak et al., 1994).Come nel caso delle CRBP, anche la struttura della oloCRABP è pressochè identica a quella della forma apo, e non è stato ancora chiarito in che modo RA sia legato o rilasciato dalla proteina (Thompson et al., 1995). Una recente ipotesi formulata da Thompson et al. (Thompson et al., 1995), basata sulla risoluzione di alcuni particolari cristalli di apoCRABP, prevede la formazione di dimeri intermolecolari aventi una struttura più “aperta” attraverso la formazione di legami idrogeno tra le catene “C” e “D” di due distinte molecole di CRABP.La funzione fisiologica della CRABP (la cui espressione è indotta da RA) non è stata ancora determinata con certezza. Una delle ipotesi è che, attraverso la formazione del complesso in forma olo, la CRABP possa regolare i livelli intracellulari di acido retinoico in forma libera disponibile per il legame con i recettori nucleari. L’espressione in tessuti distinti di due isoforme di CRABP che legano i medesimi retinoidi con differente affinità
21

Introduzione
Figura 10: Modelli di struttura della oloCRBPI (sopra) e della oloCRABPI (sotto) rappresentati evidenziando le strutture secondarie.
(la CRABPII lega 3 volte con minore affinità all-trans RA) avvalora questa ipotesi (Fiorella et al., 1993). Un possibile ruolo della oloCRABP nel processo di inattivazione dell’acido retinoico è suggerito dall’osservazione che essa è substrato, in frazioni microsomali di ratto, per la conversione catabolica dell’acido retinoico in composti più polari (Fiorella e Napoli, 1991a).E’ stato recentemente riportato da Lampron et al. (Lampron et al., 1995) il sorprendente risultato di un interessante esperimento di knock-out genico, che nega alla CRABP un ruolo essenziale: topi nei quali erano stati eliminati i geni codificanti per la CRABPI e II non sono distinguibili da animali non mutati per il loro sviluppo, la loro fertilità e longevità, e appaiono, nel loro complesso, assolutamente normali.
2.52 Esterificazione del retinolo
Esistono diversi possibili reazioni di conversione metabolica intracellulare del retinolo. Esso, all'interno delle cellule bersaglio, viene legato dalla proteina trasportatrice intracellulare, la CRBP, che lo indirizza verso specifici siti di biotrasformazione (Figura 12). E’ possibile l'accumulo sotto forma di retinil estere, oppure la mobilizzazione idrolitica mediata da distinte attività enzimatiche presenti nella frazione microsomale (Herr e Ong, 1992; Ottonello et al.,1987), tuttavia l'attivazione metabolica del retinolo mediante ossidazione ad acido retinoico rappresenta la reazione biologicamente più importante.
I retinil esteri (RE), i soli retinoidi metabolicamente inattivi e chimicamente stabili, rappresentano l’unica forma di accumulo intracellulare del retinolo, e vengono immagazzinati nelle cellule associati alle membrane del reticolo endoplasmatico.
22

Introduzione
Nelle cellule bersaglio sono presenti due distinte attività enzimatiche, presenti principalmente in frazioni microsomali di fegato o intestino, ma anche nel testicolo e nell’occhio, in grado di catalizzare il processo di esterificazione del retinolo (Blaner e Olson, 1994b).La acil-CoA retinolo aciltransferasi (ARAT) utilizza come substrati retinolo libero e acidi grassi coniugati con il Coenzima A, ma non è in grado di esterificare retinolo legato a CRBPI o II. Una seconda attività enzimatica, la cosiddetta lecitina-retinolo aciltransferasi (LRAT), impiega come donatore di gruppo acilico l’acido grasso legato al carbonio (1) della fosfatidilcolina (Mc Donald e Ong, 1988), ed è capace di esterificare in vitro sia il retinolo in forma libera che quello legato a CRBPI e II con una Km rispettivamente di 0.63, 0.20 e 0.24 M (Herr e Ong, 1992), ed una Vmax che è minore rispetto a quella
determinata per ARAT. LRAT e ARAT si differenziano anche per la diversa sensibilità ad inibitori ed allo stato nutrizionale dell’organismo. L’attività di LRAT nei microsomi epatici, a differenza di ARAT, è estremamente sensibile alla presenza nella dieta di Vitamina A, diminuendo fino a scomparire in condizioni di carenza, o incrementando notevolmente se è somministrato RA (Randolph e Ross, 1991; Matsuura e Ross, 1993).Queste caratteristiche, unitamente ai parametri cinetici che indicano quale substrato cineticamente favorito il complesso fisiologico retinolo-CRBPI o II, abbondante nel fegato o nell’intestino, rendono LRAT l’enzima presumibilmente responsabile, anche in vivo, del processo di assorbimento intestinale, e dell’immagazzinamento epatico del retinolo (Ross, 1993).Herr e Ong hanno studiato le modalità di interazione dei trasportatori intracellulari con l’enzima LRAT (Herr e Ong, 1992). E’ stato osservato un effetto differenziale di inibizione sulla reazione di esterificazione da parte della apoCRBP I e II: mentre la apoCRBPI in concentrazioni fisiologiche (dell’ordine del M) è un potente inibitore competitivo della reazione, sia in presenza di retinolo in forma libera o legato a proteine, tale effetto non è osservato nelle stesse condizioni in presenza di apoCRBPII. Questo dato, oltre a suggerire la possibilità che la concentrazione intracellulare di apoCRBPI costituisca un importante fattore per la regolazione in vivo della velocità di esterificazione, implica necessariamente che LRAT possa distinguere tra le forme apo e olo delle due proteine, riconoscendo differenze conformazionali che non sono evidenti dall’analisi della struttura nel cristallo. Alcuni dati preliminari indicano tra l’altro una possibile differenza conformazionale in soluzione tra le forme apo e olo di CRBPI e II: lo suggeriscono i loro differenti tempi di eluizione in cromatografia di scambio anionico, e la loro diversa sensibilità a proteolisi parziale impiegando differenti proteasi (Ong et al.,1994).
2.53 Idrolisi dei retinil esteri
23

Introduzione
La reazione metabolica di idrolisi dei retinil esteri riveste particolare importanza in quanto permette la mobilizzazione intracellulare del retinolo in tutti i tessuti nei quali viene accumulato. Tale attività metabolica è inoltre necessaria per i processi di l’assunzione e immagazzinamento epatico dei retinil esteri associati ai chilomicroni (Harrison, 1993).L’idrolisi dei retinil esteri è catalizzata da due distinte attività enzimatiche: una idrolasi colato-dipendente e una colato-indipendente. La prima, isolata nella frazione solubile epatica (Blaner et al.,1984), necessita di concentrazioni millimolari di colato o taurocolato per la catalisi, e possiede una scarsa specificità di substrato, in quanto è in grado di catalizzare l’idrolisi oltre che dei retinil esteri, anche di colesteril esteri e di altri trigliceridi. Le caratteristiche biochimiche e la specifica localizzazione epatica della RE idrolasi colato dipendente, rendono possibile un suo coinvolgimento nella reazione di mobilizzazione dei retinil esteri accumulati nelle cellule stellate epatiche, che costituisce la fase preliminare del processo di rilascio plasmatico del complesso retinolo-RBP (Blohomoff et al., 1985).L’attività retinil estere colato-indipendente, è stata localizzata inizialmente nella frazione microsomale epatica, ed è stata individuata anche in microsomi di reni, testicolo, intestino (Napoli et al., 1989) e occhi (Ottonello et al., 1987).Nonostante non sia stato ancora possibile purificare l’enzima responsabile dell’idrolisi colato-indipendente dei retinil esteri, le informazioni disponibili relative alle caratteristiche biochimico-funzionali, unitamente alla localizzazione intracellulare in membrana, rendono questa attività enzimatica probabilmente responsabile, in vivo, oltre che del processo di idrolisi dei RE rilasciati al fegato dai chilomicroni durante l’assorbimento, anche della mobilizzazione del retinolo dalle riserve nei vari tessuti quando le condizioni nutrizionali lo richiedono.Questa attività RE-idrolasica non necessita di sali biliari, possiede una notevole specificità di substrato, con una Km per il retinil palmitato di 50 M, e, a differenza
dall’idrolasi colato-dipendente, è sensibile alla presenza di apoCRBPI (Blaner e Olson, 1994b; Boerman e Napoli 1991). Ottonello et al. (Ottonello et al., 1987) per primi hanno individuato l’effetto stimolatorio della apoCRBPI sull’attività idrolasica colato-indipendente associata a membrane in frazioni microsomali da epitelio pigmentato di retina (RPE), il principale sito di accumulo di retinoidi nell’occhio. Questi autori hanno osservato che l’incubazione delle membrane di RPE con apoCRBPI portava all’aumento della attività idrolasica in modo dipendente dalla concentrazione della apoCRBPI, con formazione di CRBPI in forma olo come prodotto di reazione.Anche Boerman e Napoli (Boerman e Napoli, 1991) hanno osservato in frazioni microsomali isolate da fegato di ratto e da altri tessuti un analogo effetto stimolatorio della apoCRBPI. Anche in questo caso la stimolazione dell'idrolisi dei retinil esteri è dipendente dalla concentrazione della proteina in forma apo, e ha un andamento
24

Introduzione
saturabile, con il 50% della velocità massima della reazione in presenza di una concentrazione di 2.6 µM apoCRBPI, compatibile con la concentrazione endogena del trasportatore. La deplezione di substrato non rappresenta la causa primaria della saturazione, poichè in condizioni di massima velocità di reazione, non più del 10% dei retinil esteri vengono idrolizzati. Nemmeno la diminuita disponibilità di apoCRBPI è responsabile dell’andamento osservato, poichè, a saturazione, non più del 4% di essa è convertita in oloCRBPI.Nonostante la loro capacità di legare il retinolo, nè la -lattoglobulina, nè l’albumina ad una concentrazione di 90 µM hanno alcun effetto sull’idrolisi, così come l’aggiunta di CRABPI sia in forma olo che apo.L’insieme di questi dati suggerisce che la apoCRBPI possa agire mediante un’interazione diretta con l’enzima responsabile dell’attività idrolasica, piuttosto che facilitando la rimozione di retinolo dalle membrane (Boerman e Napoli, 1991).L'idrolisi dei retinil esteri, unitamente all’attivita retinolo esterasica (Vedi sopra), costituiscono le principali reazioni metaboliche responsabili del mantenimento dei livelli intracellulari di retinolo, la cui disponibilità è importante per altre reazioni metaboliche, quali ad esempio l’ossidazione ad acido retinoico. E’ importante notare l’opposto effetto modulatorio della apoCRBP, che inibisce l’immagazzinamento del retinolo nella reazione di sintesi di retinil esteri, e ne stimola invece il rilascio nella reazione inversa. La variazione dinamica della concentrazione di apoCRBP in dipendenza delle condizioni nutrizionali e dei differenti livelli di retinolo nell’organismo, potrebbe rappresentare, anche in vivo, un importante elemento di regolazione per mantenere l’omeostasi della Vitamina A, poichè, in condizioni di carenza, gli elevati livelli intracellulari di apoCRBP inibirebbero l’accumulo epatico sotto forma di retinil esteri del retinolo, e favorirebbero la sua mobilizzazione dalle riserve nel fegato e nei vari tessuti, stimolando la reazione di idrolisi, in modo opposto a quanto potrebbe accadere in condizioni di normale apporto nutrizionale.
2.54 Biosintesi di acido retinoico
La biosintesi del retinoide biologicamente più attivo è sicuramente uno dei momenti più importanti e delicati nella farmacocinetica della Vitamina A, vista l’importanza e la vastità degli effetti esercitati dall’acido retinoico. Al fine di mantenere precisi (e dinamici) livelli di concentrazione nei vari tessuti, la sintesi di acido retinoico deve essere finemente regolata, dal momento che tale molecola non è accumulabile, ed è catabolizzata rapidamente a livello intracellulare (t 1/2 = 35’; Fiorella e Napoli, 1991b). Sebbene diverse attività metaboliche potenzialmente coinvolte nel processo di sintesi dei differenti isomeri di RA biologicamente attivi siano state isolate attraverso studi in vitro, gli specifici enzimi responsabili in vivo, in condizioni fisiologiche, di tale reazione metabolica, devono ancora essere identificati. La comprensione dei meccanismi che
25

Introduzione
regolano il processo di omeostasi di RA, o l’identificazione di altri importanti effettori che modulino la sintesi, sono solo alcuni degli importanti aspetti che potranno essere chiariti solo individuando con chiarezza quali siano le vie metaboliche responsabili della sintesi di RA in condizioni fisiologiche, e isolando quindi tali attività enzimatiche.Le quantità di acido retinoico assunte con la dieta sono pressochè trascurabili. Inoltre, poichè non è mai stato evidenziato alcun organo produttore primario di acido retinoico, non è mai stato isolato un trasportatore plasmatico specifico, e i livelli plasmatici di RA presente in forma legata alla sieroalbumina sono mediamente di due ordini di grandezza più bassi di quelli misurati nei tessuti (Napoli, 1990a), è opinione corrente che esso venga prodotto in situ, attraverso l'ossidazione del retinolo, trasportato in circolo dalla RBP plasmatica e internalizzato dalle cellule bersaglio.RA costituisce uno dei metaboliti naturali (sebbene quantitativamente minoritario) del retinolo, come confermano diversi esperimenti effettuati in vivo analizzando i prodotti di conversione di ROH marcato isotopicamente, somministrato in dosi fisiologiche con la dieta. La capacità di convertire retinolo, fornito in forma libera, in acido retinoico è stata dimostrata in numerosi tessuti o in colture cellulari di differenti specie animali (uomo, cane, gatto, ratto, topo, maiale) (Napoli, 1994).Diversi studi in vitro utilizzanti estratti cellulari di differenti tessuti e specie animali, hanno tentato di identificare ed isolare le attività enzimatiche responsabili della sintesi di acido retinoico da retinolo in forma libera, ma con risultati spesso contrastanti riguardo la possibile identificazione dell’attività responsabile della biosintesi di RA. La frazione citosolica di vari tessuti, ricca di alcool e di aldeide deidrogenasi, è in grado di sintetizzare acido retinoico, in presenza di NAD come cofattore, da retinolo libero disperso in solvente organico o in detergenti, con accumulo in quantità minima dell’intermedio retinaldeide (Napoli, 1994; Napoli e Race, 1987; Napoli e Race, 1990b; Posch et al., 1989; Siegenthaler et al., 1990). Tale sintesi è risultata in alcuni casi essere catalizzata da alcool deidrogenasi (ADH) normalmente coinvolte nel metabolismo dell'etanolo, oppure, in altri casi, da retinolo deidrogenasi distinte dalle precedenti, in presenza della frazione solubile di vari tessuti, ricca di aldeide deidrogenasi (ALDH) (Kim et al., 1992; Posch e Napoli, 1992b).Il principale prodotto di incubazione quando è impiegato retinolo in forma libera quale substrato in presenza della frazione microsomale è invece retinaldeide, e, solo in minima parte, RA. Infatti i microsomi, nei quali si localizzano attività enzimatiche coinvolte nel catabolismo della Vitamina A oltre a quelle di sintesi e mobilizzazione di retinil esteri, possiedono distinte attività NAD e NADP-dipendenti che convertono retinolo a retinaldeide, ma non sono in grado di ossidare ulteriormente quest’ultima in modo efficiente (Napoli e Race, 1987; Leo et al., 1987; Napoli et al., 1992). Un’altra interessante caratteristica dei microsomi è che essi interferiscono drasticamente con la sintesi citosolica di acido retinoico (Posch et al., 1989; Napoli e Race, 1990b; Kim et al., 1992).
26

Introduzione
Sebbene siano state isolate diverse attività enzimatiche distinte dalle ADH in grado di ossidare il retinolo a retinaldeide tra le quali una RoDH microsomale in fegato di ratto (34 KDa), appartenente alla famiglia delle SCAD (short-chain alcohol dehydrogenase) (Vedi oltre), il possibile ruolo delle alcool deidrogenasi nella sintesi in vivo di RA è ancora oggi discusso.Le alcool deidrogenasi citosoliche coinvolte nel metabolismo dell’etanolo, presenti come isoenzimi in uomo, ratto e topo, sono raggruppate in quattro classi, distinte per caratteristiche biochimiche e funzionali (Tabella 3), e sono presenti in forma cataliticamente attiva prevalentemente come dimeri, costituiti da subunità aventi peso molecolare di circa 35-40000 (Jörnvall et al.,1993).Ad eccezione delle ADH aventi caratteristiche analoghe a quelle umane di classe III, che sono coinvolte nell’ossidazione della formaldeide in presenza di glutatione, le altre classi di ADH sono tutte in grado di ossidare in vitro il retinolo a retinaldeide in presenza di NAD, sebbene con differente efficienza catalitica. Queste attività sono classificabili per la loro specifica localizzazione tissutale e mostrano differente sensibilità ad uno specifico inibitore, il pirazolo (Yang et al., 1994). Esperimenti effettuati in vitro in differenti animali hanno mostrato che soprattutto le alcool deidrogenasi analoghe alle classi I e IV potrebbero possedere i requisiti necessari per svolgere un possibile ruolo nella biosintesi di RA.
Uomo Localizzazione Caratteristiche funzionali Ratto Topo
Classe I (ADH1, 2, 3)
Fegato, occhi, intestino
Km ROH » 50 M (1); 35M (2); 290M
(3) Elevata sensib. pirazolo (Ki: 1 M)
ADH-3 ADH-A2
Classe II (ADH2)
Fegato Media sensib. pirazolo
Km ROH » 14 M- -
Classe III (ADH3)
Ubiquitaria Non ossida retinoloInsensibile al pirazolo (Ki: 80 mM)
ADH-2 ADH-B2
Classe IV (ADH4)
Stomaco Km ROH » 10 M Media sensib. pirazolo (Ki: 0.6-2 mM)
ADH-1 ADH-C2
Tabella 3: Classificazione delle ADH secondo criteri biochimico-funzionali.
La classe I delle ADH umane è responsabile, in vivo, dell’ossidazione dell’etanolo nel fegato, organo nel quale sono espresse in prevalenza, mentre in ratto e topo gli enzimi analoghi sono abbondanti anche nel tessuto oculare e nell’intestino. Gli isoenzimi di questa classe sono quelli che presentano la maggiore sensibilità al pirazolo, con una Ki di circa 1 mM. La recente scoperta di un elemento di risposta all’acido retinoico (RARE) a monte del gene umano codificante per la ADH3 umana citosolica di classe I, è sicuramente un dato interessante, anche se non implica necessariamente che tale enzima
27

Introduzione
debba essere coinvolto proprio nel processo di biosintesi di RA (Duester et al., 1991). Tra l’altro, ADH3 è quello che presenta i parametri cinetici (Km, e Vmax) meno favorevoli nel catalizzare l’ossidazione del retinolo, rispetto agli altri isoenzimi appartenenti alla classe I delle ADH (Tabella 3; Yang et al., 1994).La classe IV delle ADH umane risulta espressa esclusivamente nello stomaco, ma gli enzimi analoghi di ratto e topo sono abbondanti anche negli epiteli e nel tessuto oculare. Questa classe di alcool deidrogenasi possiede una sensibilità intermedia al pirazolo (Ki: 0.6-2 mM), e possiede il miglior rapporto tra Vmax e Km per l’ossidazione del retinolo.La retinaldeide sintetizzata da retinolo deidrogenasi microsomali, o da alcool deidrogenasi citosoliche, deve essere ossidata ulteriormente per formare acido retinoico. Anche in questo caso, come per le ADH, differenti aldeide deidrogenasi che risultano in grado di catalizzare in vitro l’ossidazione della RALD a RA sono state identificate nell’uomo o in altre specie animali, ma senza ottenere risultati univoci riguardo l’individuazione dell’enzima in grado di catalizzare in vivo questa reazione. Nella maggior parte dei casi, le aldeide deidrogenasi identificate o purificate, hanno un peso molecolare di circa 200000-230000, e risultano costituite da tetrameri di 55 kDa (Sladek et al., 1989).Nell’uomo esistono tre classi di ALDH, che si distinguono per la localizzazione tissutale e per la specificità di substrato: le ALDH1 e 2 sono presenti principalmente nel fegato, rispettivamente nel citoplasma e nei mitocondri, mentre la ALDH3 è espressa esclusivamente nella frazione solubile dello stomaco. Tra queste, solamente la ALDH1 è in grado di catalizzare in vitro la sintesi NAD-dipendente di RA da retinaldeide, tra l’altro con una elevata efficienza catalitica (Km: 60 nM) nel caso dell’enzima
ricombinante (Yoshida et al., 1994). L’enzima corrispondente nel topo, alla ALDH1 umana, è la ALDH-2, presente nel citoplasma di fegato, intestino e organi genitali, che è in grado di ossidare la retinaldeide in frazioni subcellulari con una Km minore di 1 M (Lee et al., 1991).
Nel ratto sono state isolate aldeide deidrogenasi in fegato e reni, simili alla ALDH1 umana per punto isoelettrico (pH 8.3) e per la capacità di ossidare la retinaldeide (Posch et al.,1992a; Labrecque et al., 1993). L’enzima purificato da fegato di ratto è una retinaldeide deidrogenasi avente P.M. apparente di 220000 (subunità di 55 kDa) che ossida RALD presente in concentrazioni M (Vedi sistema “misto”), mentre l’attività isolata da reni presenta una Km di 8 M per la retinaldeide, ed ha peso molecolare apparente di 140000 (subunità di 53 kDa). Una retinaldeide deidrogenasi abbondante in retina bovina (Km: 9 M) è stata recentemente purificata e sequenziata: è costituita da subunità di 55 kDa, e presenta l’88% di identità di sequenza amminoacidica con la ALDH1 umana (Saari et al., 1995).Nonostante queste attività enzimatiche, citosoliche e microsomali, siano in grado di catalizzare in vitro l’ossidazione di concentrazioni micromolari di retinolo e retinaldeide in forma libera, le condizioni notevolmente differenti presenti in vivo suggeriscono
28

Introduzione
moderazione nell’attribuire ad enzimi appartenenti alla famiglia delle ADH e delle ALDH un ruolo importante nella biosintesi di RA: la concentrazione di retinoidi impiegata come substrato è superiore a quella fisiologica, ed i valori di Km riferibili a tali
attività sono ampiamente superiori alla concentrazione intracellulare di retinolo libero. Infatti, la formazione del complesso retinolo-CRBP, forma principale nella quale il retinolo si trova disponibile in vivo, limita notevolemente la quantità di retinolo in equilibrio, prevenendone presumibilmente l’ossidazione da parte attività enzimatiche aspecifiche.Il complesso retinolo-CRBP, la cui concentrazione intracellulare stimata è di circa 6-7 M (Yost et al., 1988), rappresenta presumibilmente il substrato fisiologicamente significativo per la sintesi in vivo di acido retinoico. La presenza nelle cellule di un eccesso di apo-proteina (Harrison et al., 1987) e la bassa costante di dissociazione del complesso retinolo-CRBP supportano questa ipotesi, sebbene non sia stata ancora ottenuta alcuna evidenza sperimentale diretta dell’interazione tra CRBP ed enzima ossidante. Tuttavia, diversi dati riportati in letteratura (vedi esterificazione del retinolo e idrolisi degli esteri) hanno evidenziato uno specifico ruolo della CRBP (in forma olo e apo) nel veicolare il retinolo verso specifici siti di metabolizzazione intracellulare e nel modulare attività enzimatiche (Ottonello et al., 1987; Boerman e Napoli, 1991; Herr e Ong, 1992).Due distinte vie biosintetiche capaci di sintetizzare acido retinoico in vitro utilizzando quale substrato il complesso all-trans retinolo-CRBPI in concentrazioni fisiologiche quale substrato sono state identificate negli ultimi anni in frazioni subcellulari di ratto e bue.
2.55 Il sistema “misto”
Una delle due vie di sintesi, il cosiddetto sistema “misto”, la cui presenza è stata rilevata da Posch et al. inizialmente in frazioni subcellulari di fegato di ratto (Posch et al., 1991; Posch et al., 1992a), prevede una reazione a due step che richiede sia la presenza di una frazione microsomale che citosolica. Nella prima è presente una deidrogenasi NADP-dipendente che catalizza l’ossidazione del retinolo legato alla CRBP a retinaldeide, la quale, nella frazione solubile, è substrato per la sintesi di acido retinoico da parte di una deidrogenasi citosolica NAD-dipendente.Il primo enzima è una retinolo-deidrogenasi (RoDH; 34 KDa), che è stata recentemente clonata, espressa e caratterizzata, ed è appartenente alla famiglia delle short-chain alcohol dehydrogenase (SCAD)(Boerman e Napoli, 1995; Chai et al., 1995). Nei saggi in vitro, la RoDH catalizza l’ossidazione NADP-dipendente di all-trans retinolo, in forma libera o legato a CRBP, a retinaldeide, con parametri farmacocinetici (Tabella 4) compatibili con la concentrazione fisiologica del complesso retinolo-CRBP, ma non con quella intracellulare del retinolo in equilibrio. Infatti la concentrazione
29

Introduzione
stimata di quest’ultimo è di almeno due ordini di grandezza inferiore alla Km determinata
per l’ossidazione del substrato in forma libera.
Substrato Km Vmax
(pmol/h mg di proteina)
OloCRBPI 1.9 1380
Retinolo 4 7860
Tabella 4: Parametri cinetici relativi alla sintesi di retinaldeide in microsomi di ratto in presenza di retinolo in forma libera o legato a CRBPI, con NADP come coenzima (Da Napoli, 1994).
Esperimenti in cui era rilevata la sintesi di retinaldeide da parte della RoDH, mantenendo costante la concentrazione di ROH libero e variando quella di oloCRBP, o viceversa, hanno evidenziato come fosse presente un riconoscimento preferenziale da parte dell’attività enzimatica ossidasica del complesso retinolo-CRBP, piuttosto che del retinolo presente in equilibrio.L’attività solubile del sistema “misto” è costituita da una retinaldeide deidrogenasi tetramerica NAD-dipendente (RalDH), che è stata purificata da citosol di fegato di ratto più di 2000 volte (Posch et al., 1992a). La RalDH ha evidenziato la capacità di poter utilizzare come substrato in vitro sia retinaldeide in forma libera che incubata con un eccesso di apoCRBPI, (Posch et al., 1992a). Anche in questo caso, esperimenti analoghi a quelli appena descritti per la RoDH, sembrano evidenziare un riconoscimento preferito del substrato nella forma legata, ipotizzando per la prima volta un ruolo fisiologico per il complesso retinaldeide-CRBPI, ma occorre tenere presente che è mai stata riportata alcuna evidenza dell’esistenza in vivo di tale complesso, come d’altra parte della retinaldeide in forma libera, al di fuori del tessuto oculare (Wolf, 1984). Sembra che la RalDH possieda una buona selettività rispetto alla configurazione isomerica del substrato: è in grado di ossidare gli isomeri all-trans e 9-cis, ma non riconosce la 13-cis retinaldeide. La presenza di tale attività enzimatica è stata rilevata oltre che nel fegato, in altri organi di ratto, quali rene e testicolo. Secondo il modello proposto da Napoli i due step ossidativi necessari per la sintesi di RA avverrebbero senza la diffusione dei retinoidi nel mezzo di reazione e sarebbero entrambi mediati da specifici riconoscimenti proteina-proteina (CRBP-deidrogenasi). Non è stato ancora chiarito se, e in che modo, sia presente una qualche forma di riconoscimento, o una specifica associazione funzionale tra le due deidrogenasi, localizzate in compartimenti subcellulari distinti, e in quale forma l’intermedio retinaldeide venga trasportato dai microsomi al citosol. Una ipotesi, non supportata da alcuna evidenza sperimentale diretta, è che la retinaldeide generata nel corso del primo step ossidativo rimanga legata alla apoCRBP, che fungerebbe così da shuttle per presentare poi la retinaldeide alla deidrogenasi citosolica (Posch et al., 1992a).
30

Introduzione
Nonostante siano state ottenute evidenze sperimentali che supportano l’esistenza di uno specifico riconoscimento del complesso retinolo-CRBP da parte delle attività ossidasiche RoDH e RalDH del sistema “misto”, queste, in forma parzialmente purificata, oppure espresse in cellule eucariote in seguito a clonaggio, possiedono sempre la capacità di riconoscere efficientemente quale substrato anche retinolo o retinaldeide in forma libera, con Km e Vmax più elevate (Boerman e Napoli, 1995; Napoli, 1994). Questo dato
evidenzia la presenza di una selettività non assoluta da parte del sistema ossidante per i substrati legati a CRBP, e suggerisce che il rilascio del ligando dalla proteina sia l’evento limitante per la catalisi, rendendo quindi l’impiego dei retinoidi dispersi in solvente organico comunque possibile per vari tipi di analisi.
2.56 Il sistema “solubile”
Una differente via enzimatica per la sintesi di RA da oloCRBPI che, a differenza della precedente, necessita della sola frazione citosolica, è il cosiddetto “sistema solubile”, identificato per la prima volta nel nostro laboratorio in estratti da fegato bovino (Ottonello et al., 1993).La reazione di sintesi in vitro di RA prevede l’impiego del complesso [3H] retinolo-CRBPI quale substrato, procede in presenza dei cofattori NAD o NADP ad un pH ottimale di 8, senza accumulo apparente dell’intermedio di reazione retinaldeide (suggerendo così che la deidrogenazione del retinolo sia il passaggio limitante del processo ossidativo), ed è stata rilevata anche nel citosol di altri tessuti bovini quali rene, testicolo e epitelio pigmentato, sebbene l’attività ossidasica sia minore di quella determinata nel fegato.La sintesi citosolica di acido retinoico è saturabile, dipendente dalla concentrazione di oloCRBPI, piuttosto che dal retinolo in equilibrio, e presenta parametri cinetici (Km: 0.75 M e Vmax: 625 pmoli/ h mg di proteina) fisiologicamente significativi (compatibili con
la concentrazione intracellulare della CRBPI e di RA).Il retinolo in forma libera, disperso in dimetilsolfossido (DMSO), è ossidato ad acido retinoico dal citosol di fegato bovino, con una Km di 7 M, ed una Vmax di 950 pmol/h
mg di proteina. L’effetto del sovente organico DMSO sulla sintesi di RA da oloCRBPI è quello di elevare la Km di 4 volte, senza alterare la Vmax. I parametri farmacocinetici
determinati, assumendo un effetto lineare del solvente organico, e considerato il valore della costante di dissociazione del retinolo dalla CRBPI, suggeriscono che il complesso retinolo-CRBP, piuttosto che il retinolo in equilibrio possa essere l’effettivo substrato della reazione. La sintesi di RA sia da retinolo libero che in forma legata è pressochè insensibile alla presenza di 1mM pirazolo, uno specifico inibitore delle alcooldeidrogenasi, il che distingue il sistema enzimatico “solubile” che ossida la oloCRBP in
31

Introduzione
Title: D:\CACCA\FIG00.PSCreator: GSS*CGI PostScript DriverCreationDate: 4 February 1996 17:37
Figura 11: Effetto della apo-CRBPI e di altre retinoid-binding proteins bovine sulla sintesi di RA da [3H] retinolo-CRBP. Quantità crescenti di apoCRBPI bovina ( l ), apoCRABPI ( u ), apo RBP ( o ), -lattoglobulina ( ), e sieroalbumina ( n ) sono aggiunte alla miscela di reazione contenente 1 M [3H] retinolo-CRBP. La curva tratteggiata rappresenta la concentrazione stimata di retinolo in equilibrio con la oloCRBPI alle varie concentrazioni di apoCRBPI; il 100% corrisponde in questo caso ad una concentrazione di retinolo in equilibrio di 0.1 M (da Ottonello et al., 1993)
estratti da fegato bovino dalle ADH umane di classe I e II. La apoCRBPI si è dimostrata essere un potente inibitore della sintesi solubile da oloCRBPI (Figura 11). L’effetto osservato (55% inibizione in presenza di concentrazioni equimolari di apoCRBPI) sembra essere specifico per la CRBPI, poichè altre apo-proteine leganti il retinolo quali -lattoglobulina, sieroalbumina, o RBP, oltre alla apoCRABP, proteina strutturalmente omologa alla CRBPI, non inibiscono significativamente la reazione. Il profilo di inibizione osservato in presenza di concentrazioni crescenti di apoCRBPI non correla con il calo teorico di retinolo libero in equilibrio con la oloCRBPI substrato, cosicché l’effetto osservato non è spiegabile semplicemente con il sequestramento del retinolo in equilibrio, ma rende invece possibile l’esistenza di un riconoscimento diretto tra la apoCRBP e l’attività ossidante.La sintesi “solubile” di RA a partire da [3H] retinolo-CRBP sembra avvenire mediante un meccanismo di trasferimento diretto dell'intermedio retinaldeide all'interno del sistema enzimatico di ossidazione, che ne impedisce il rilascio e l'equilibramento nel mezzo di reazione, in quanto non è influenzata dalla presenza di elevate concentrazioni (30 M) di retinaldeide non marcata radioattivamente.
32

Introduzione
?
DNA
9-cis RA
RAR (, )
RXR (, )
All-trans RA
NUCLEO
CITOPLASMA
9-cis RA
All-trans retinolo
CRBPI
Retinil esteri
LRAT ARAT
REH
apo CRBPI
All-trans RA
Figura 12: Schema del metabolismo intracellulare della Vitamina A.
33

Finalità della Ricerca
3. FINALITA' DELLA RICERCA
Considerando la molteplicità delle azioni esercitate e la complessità del metabolismo dei retinoidi, è ragionevole ritenere che la sintesi regolata di acido retinoico possa svolgere un ruolo assolutamente centrale nel controllo di processi biologicamente importanti quali la regolazione della crescita e il differenziamento cellulare, con importanti implicazioni a livello farmacologico.Il presente studio si è proposto quindi di chiarire ed approfondire differenti aspetti del processo di attivazione intracellulare del retinolo ad acido retinoico, evento importante per la farmacocinetica della vitamina A. Gli obiettivi principali verso i quali è stato rivolto il lavoro di ricerca sono stati i seguenti:
· Ottimizzazione del microsaggio per la rivelazione dell’attività retinolo ossidasica al fine di permetterne l’applicazione a tipi cellulari per i quali la sintesi di acido retinoico può rivestire interesse sia diagnostico che terapeutico
· Isolamento, caratterizzazione e parziale purificazione del sistema enzimatico retinoico biosintetico presente nel citosol di fegato bovino
· Caratterizzazione e ottimizzazione biochimica delle due vie di sintesi di acido retinoico in frazioni subcellulari da fegato di ratto; confronto con l’attività acido retinoico biosintetica precedentemente identificata in estratti epatici bovini
· Verifica sperimentale di possibili vie di formazione dell’acido 9-cis retinoico: isomerizzazione post-sintetica, origine metabolica da all trans retinolo-CRBP, o da precursori aventi configurazione isomerica 9-cis
· Proprietà biochimico-funzionali del complesso 9-cis retinolo-CRBP· Analisi della configurazione isomerica dell'acido retinoico prodotto dai due sistemi
biosintetici, "solubile" e "misto", in frazioni subcellulari da tessuti di ratto e bue utilizzando quale substrato il complesso fisiologico all trans e 9-cis retinolo-CRBP.
· Reattività dei due sistemi biosintetici nei confronti di differenti forme isomeriche di retinolo in forma libera.
· Effetto dei trasportatori intracellulari sul processo di idrolisi dei retinil esteri; analisi della configurazione isomerica del retinolo mobilizzato nella reazione retinil estere idrolasica.
· Analisi della configurazione isomerica e della reattività del retinolo endogeno presente in frazioni subcellulari da fegato di ratto e bue.
· Parziale purificazione, identificazione ed analisi funzionale dei componenti proteici responsabili della sintesi del sistema "solubile" in citosol da fegato di ratto
· Impiego del saggio di biosintesi in vitro per la caratterizzazione biochimica della formazione di acido retinoico in vari tipi cellulari del testicolo di ratto, un modello sperimentale strettamente dipendente dalla sintesi endogena di acido retinoico
34

Risultati
4. RISULTATI
4.1 Ottimizzazione del saggio in vitro per la sintesi di acido retinoico
Al fine di rendere più rapido, preciso e versatile il saggio in vitro per la sintesi di acido retinoico, e di consentirne l’impiego per l’analisi della capacità RA biosintetica di estratti cellulari provenienti da qualsiasi tessuto animale, sono state apportate diverse modifiche alla procedura precedentemente impiegata, basata sull’uso di [3H] retinolo-CRBPI purificata da organi bovini.Il nuovo sistema di dosaggio utilizza invece come substrato oloCRBPI di ratto non marcata radioattivamente, ottenibile mediante la tecnologia del DNA ricombinante in forma pressochè pura ed in elevate quantità. E’ stato inoltre aumentato il volume di reazione da 50 a 200 l, ottimizzato il processo di estrazione in fase organica dei retinoidi, ed approntata una migliorata analisi dei prodotti in HPLC. Tutte queste modifiche ci hanno consentito di allestire un sistema di dosaggio che, rispetto a quello precedente, presenta una sensibilità di rivelazione (1 pmole) comunque notevole, utilizza substrati meno costosi e pericolosi, e permette un’analisi diretta ed estremamente rapida dei prodotti di reazione (Figura 13).
Incubazione per 30’a 37 °C
Analisi in HPLC deiretinoidi
Estrazione dei retinoidiin solvente organico
Elaborazione dei dati
Quantità (pmoli) diprodotti sintetizzati
·Olo-CRBPI +/- apoCRBPricombinante di ratto
·NAD+ (e NADP+ nel sistema“misto”)
·Citosol di fegato bovinoo di ratto
·Microsomi di fegato bovino odi ratto (nel sistema “misto”)
·Buffer, inibitori di proteasi
Figura 13: Le fasi del saggio “a freddo” di biosintesi di acido retinoico.
35

Risultati
L’impiego quale substrato di una proteina eterologa (CRBPI di ratto), pur se dotata di una notevole omologia di sequenza (97%) con la corrispondente proteina bovina, ha richiesto il confronto dei parametri cinetici della reazione di sintesi NAD-dipendente di RA con quelli precedentemente ottenuti utilizzando quale substrato la proteina omologa.
Title: C:\CAVA\FIG02.EPSCreator: GSS*CGI PostScript DriverCreationDate: 13 February 1996 16:29
Figura 14: (A) Dipendenza della sintesi citosolica di RA da concentrazioni crescenti di [ 3H] retinolo-CRBPI bovina ( ) o oloCRBPI ricombinante di ratto ( ), in presenza di NAD come cofattore.
Substrato CoenzimaKm
(M)
Vmax
(pmol/ h mg proteina)
Retinolo-CRBPI di ratto NAD 1.7 514
Retinolo-CRBPI di ratto NADP 1.3 200
[3H] retinolo-CRBPI bovina NAD 0.8 601
[3H] retinolo (2% DMSO) NAD 7.1 948
[3H] retinolo-CRBPI bovina (2% DMSO) NAD 3.5 611
Tabella 5: Parametri cinetici relativi alla sintesi di RA in frazioni citosoliche bovine da retinolo disperso in solvente organico, oppure legato a CRBPI bovina o ricombinante di ratto. Sono state impiegate condizioni standard di reazione (200 l di volume, incubazione per 30’ a 37°C, pH 8). Le reazioni sono state eseguite in presenza di 200 g di proteina citosolica, con 2 mM NAD o NADP come cofattore. I parametri cinetici qui riportati quando è impiegato come substrato [3H] retinolo disperso in DMSO o legato alla CRBPI, sono quelli ottenuti da Ottonello et al. (Ottonello et al., 1993), con il precedente saggio di attività RA biosintetica.
36

Risultati
La curva di saturazione relativa alla sintesi di RA in citosol di bue, ottenuta utilizzando concentrazioni crescenti di oloCRBP ricombinante di ratto come substrato, è analoga a quella precedentemente osservata impiegando [3H] retinolo-CRBPI bovina, così come sono simili i parametri cinetici relativi (Figura 14; Tabella 5).Si è inoltre confermato, utilizzando il saggio “a freddo” con oloCRBPI di ratto come substrato, l’effetto inibitorio della apoCRBPI sulla sintesi di acido retinoico. In questo caso l’ottima sovrapponibilità della curva di inibizione ottenuta precedentemente con proteine bovine rispetto a quella ricavata utilizzando CRBPI ricombinante di ratto (dato non mostrato), conferma l’ipotesi di un possibile riconoscimento CRBP-sistema ossidante per spiegare il fenomeno, considerato che la Kdiss del complesso retinolo-
CRBP di ratto è di circa 10-10 M (Li et al.,1991).
4.2 Il sistema “misto” in estratti subcellulari da fegato bovino
L’identificazione del cosiddetto sistema biosintetico “misto” in organi di ratto da parte di Posch et al. (Posch et al. 1992a) ci ha indotto a verificare la presenza di tale attività in frazioni subcellulari di fegato bovino. Impiegando il nuovo saggio “a freddo” con oloCRBPI ricombinante di ratto quale substrato, è stato possibile rilevare per la prima volta l’attività biosintetica di tale sistema anche in frazioni subcellulari microsomali e citosoliche di fegato bovino, che risultano in grado di sintetizzare acido retinoico con una velocità circa 6 volte superiore rispetto a quella del sistema citosolico (Ottonello et al., 1994). I parametri cinetici relativi al primo step ossidativo del sistema “misto”, la sintesi microsomale di retinaldeide, sono stati determinati, ed è stato confermato l’uso del NADP come cofattore, con valori di Km e Vmax che sono in buon accordo con quelli
precedentemente determinati in ratto (cfr. Tabella 6 e 4).
Substrato CoenzimaKm
(M)
Vmax (pmol/ h mg proteina)
Retinolo-CRBPI di ratto NADP 2.2 3630
Retinolo-CRBPI di ratto NAD 1.2 602
Tabella 6: Parametri cinetici relativi alla sintesi di retinaldeide in frazioni microsomali bovine da retinolo-CRBPI ricombinante di ratto. Le reazioni sono state eseguite in condizioni standard di reazione (200 l di volume, incubazione di 30’ a 37°C, pH 8), in presenza di 200 g di proteina microsomale, con 2 mM NAD o NADP come cofattore .
Al fine di individuare il rapporto ottimale tra proteina citosolica e microsomale necessario per ottenere la massima sintesi di acido retinoico nel sistema “misto” di fegato bovino, è stata rilevata la formazione di RA e l’accumulo di retinaldeide generata nella
37

Risultati
frazione microsomale, non convertita dalla deidrogenasi NAD-dipendente solubile variando il rapporto tra microsomi e citosol (Figura 15).
Title: D:\CACCA\FIG03.PSCreator: GSS*CGI PostScript DriverCreationDate: 4 February 1996 17:47
Figura 15: Sintesi di RA da oloCRBPI di ratto in presenza di una quantità fissa di proteina citosolica, e quantità crescenti di microsomi isolati da fegato bovino. A 200 g di citosol sono aggiunti fino a 400 g di microsomi, in presenza di 2 mM NAD e NADP e 5 M olo CRBPI di ratto.
La sintesi di RA nel sistema “misto” bovino in presenza di una quantità costante di proteina citosolica e crescente di proteina microsomale ha mostrato un massimo di attività ad un rapporto tra le due frazioni di circa 1:1. In queste condizioni di reazione, l’intermedio di reazione retinaldeide è chiaramente rilevabile, e tende ad accumularsi con l’aumentare della quantità di microsomi impiegati.
4.3 Parziale purificazione dell’attività enzimatica solubile da citosol di fegato bovino
Il preliminare isolamento del sistema biosintetico solubile da citosol di fegato bovino è stato ottenuto frazionando il citosol su di un gradiente di glicerolo (15-30%) in presenza di una elevata concentrazione salina (500 mM KCl). Le frazioni così ottenute sono state testate con il saggio di attività retinoico biosintetica per il sistema “solubile”, che ha mostrato un picco di attività NAD-dipendente nella regione avente un peso molecolare apparente di circa 130 kDa, distinto da quello della maggior parte delle alcool deidrogenasi (Figura 16, A). Così come nel citosol non frazionato, la reazione è in grado di impiegare oloCRBPI di ratto quale substrato, e procede senza accumulo di retinaldeide nella frazione di massima sintesi. Un picco distinto di attività retinoico biosintetica viene
38

Risultati
identificato testando le frazioni del gradiente per la capacità di ossidare la retinaldeide
Title: D:\CACCA\FIG04.PSCreator: GSS*CGI PostScript DriverCreationDate: 4 February 1996 17:52
Figura 16: Frazionamento in gradiente di glicerolo di citosol da fegato bovino. Uguali volumi delle frazioni raccolte sono stati testati per la sintesi di RA ( l ) in assenza (A) o in presenza (B) di un eccesso di microsomi. La concentrazione di oloCRBPI è di 5 M, e 2 mM NAD in (A) o 2 mM ciascuno NAD e NADP (B) sono presenti come cofattori. La quantità di RA sintetizzato nelle reazioni con le frazioni 8 (A) e 12 (B) è rispettivamente di 35 e 330 pmoli. Il 100% di retinaldeide non convertita ( ¡ ) in (B) corrisponde a 315 pmoli.
39

Risultati
generata da retinolo-CRBPI in presenza di frazioni microsomali (Figura 16, B). Il peso molecolare apparente della retinaldeide deidrogenasi del sistema “misto” di bue così identificata, circa 230 kDa, è in buon accordo con i dati di Posch et al. (Posch et al, 1992a), ed è ben distinto da quello della retinolo deidrogenasi del sistema “solubile”.I due picchi di attività RA biosintetica identificati sono distinti anche funzionalmente: non si osserva un aumento significativo della sintesi di RA aggiungendo microsomi alla frazione di 130 kDa, così come la frazione di 230 kDa non è in grado di ossidare retinolo-CRBP ad acido retinoico.Per verificare la possibilità che il componente ”solubile” di 130 kDa fosse in realtà costituito dall’intersezione di due distinti componenti enzimatici, le due frazioni laterali del picco sono state riunite e testate per l’attività RA biosintetica, che è risultata essere additiva, e non sinergica. Nei limiti della risoluzione di questo tipo di analisi in gradiente di glicerolo, il picco di 130 kDa non sembra quindi essere il prodotto dell’intersezione di un’alcool e un’aldeide deidrogenasi. Nonostante il componente solubile del sistema “misto” mostri un arricchimento di circa 6 volte rispetto all’attività specifica del citosol non frazionato, il recupero quantitativo dell’attività solubile è piuttosto basso (meno del 30%), e la sua attività specifica non è maggiore di quella del citosol non frazionato; la causa della scarsa purificazione ottenuta non è stata identificata con certezza, ma potrebbe essere imputabile alla notevole labilità dell’attività solubile.Sono stati effettuati vari tentativi per isolare e purificare l’attività enzimatica “solubile” presente in citosol di fegato bovino: una prova preliminare di frazionamento selettivo in presenza di differenti percentuali di solfato d’ammonio non ha portato ad un incremento significativo dell’attività specifica. Diversi metodi cromatografici standard sono stati impiegati successivamente, quali colonne a scambio anionico e cationico deboli e forti, di idrossiapatite, matrici di affinità contenenti un analogo del NAD: nonostante siano state impiegate in alcuni casi per la stessa colonna differenti condizioni di eluizione, e, nel caso della colonna a scambio cationico forte e idrossiapatite, l’attività retinoico biosintetica sia stata inizialmente trattenuta dalla matrice cromatografica, si è sempre ottenuto un basso recupero quantitativo, e soprattutto, non è stato rilevato nessun incremento dell’attività specifica che potesse far pensare di aver individuato condizioni favorevoli per un arricchimento consistente dell’attività RA biosintetica solubile. E’ possibile che le condizioni di eluizione impiegate (pH acido o basico, elevate concentrazioni saline) abbiano causato la parziale inattivazione dell’attività enzimatica solubile. La scarsa attività recuperata potrebbe anche essere stata provocata dalla rimozione di effettori endogeni importanti per la catalisi, oppure dalla dissociazione dei componenti proteici dell’apparato enzimatico responsabile della sintesi di RA a partire da olo-CRBPI di ratto.Un differente approccio cromatografico è stato impiegato al fine di ottenere un significativo incremento dell’attività specifica, ma soprattutto per dimostrare l’esistenza di una interazione diretta tra il sistema enzimatico ed il complesso retinolo-CRBPI,
40

Risultati
ipotizzata sulla base dei dati disponibili. Avvantaggiandoci della possibilità di ottenere CRBPI di ratto in grandi quantità grazie alla tecnologia del DNA ricombinante, abbiamo allestito una matrice di affinità biospecifica contenente CRBP di ratto in forma immobilizzata (sia apo che olo), e, come controllo una colonna con sieroalbumina. Purtroppo anche in questo caso non sono state identificate condizioni di legame che consentissero una interazione stabile fra la CRBPI e l’attività enzimatica solubile di 130 kDa: l’attività presente nel citosol bovino caricato sulla colonna di affinità contenente CRBPI è stata recuperata sempre nell’eluato, in modo perfettamente sovrapponibile a quanto avveniva per la colonna con sieroalbumina. Nemmeno impiegando la frazione arricchita con la retinaldeide deidrogenasi di 220 kDa si sono ottenuti risultati differenti. Verosimilmente, l’interazione tra la CRBPI e i sistemi ossidativi solubili non è abbastanza forte e duratura per poter essere evidenziata con questo tipo di matrice di affinità.
4.4 Identificazione e caratterizzazione delle vie di biosintesi di RA in estratti cellulari da fegato di ratto. Confronto con l’attività identificata in organi bovini
Avendo identificato l’esistenza di entrambe le vie, “solubile” e “mista”, di sintesi di RA da oloCRBPI in estratti cellulari da fegato bovino, e avendo tentato, senza successo, di purificare l’attività del sistema “solubile”, ci siamo preoccupati di verificare l’esistenza e caratterizzare tali attività anche in frazioni subcellulari da fegato di ratto, analizzarne le caratteristiche biochimiche e funzionali, ed effettuare eventualmente un nuovo tentativo di isolamento del sistema enzimatico.Come mostrato in Tabella 7, applicando il microsaggio in vitro per testare l’attività ossidante “solubile” e “mista” con all-trans retinolo-CRBPI di ratto in concentrazioni fisiologiche (4 M), in presenza di frazioni subcellulari preparate da fegato di ratti adulti,
Specie Sistema enzimatico Acido retinoico (pmoli)
oloCRBPI oloCRBPI + apoCRBPI
Bue Solubile 36 17
Bue Misto 310 170
Ratto Solubile 35 16
Ratto Misto 34 17
Lievito Solubile N.D. N.D.
Lievito Mistoa N.D. N.D.
Tabella 7: Sintesi di acido retinoico in differenti frazioni subcellulari di bue, ratto e lievito. Sono state impiegate condizioni standard di reazione (30’, 37°C, 2 mM NAD e/o NADP), utilizzando 200 g di
41

Risultati
proteina citosolica o 200+200 g di proteina microsomale nel caso del sistema “misto”; la concentrazione di olo o apoCRBPI è di 4 M. aIn questo caso sono state incubate 200 g di proteina microsomale di bue o ratto con 200 g di proteina citosolica di lievito.
l’attività di entrambe le vie di sintesi è chiaramente rilevabile. Confrontando i valori di attività con quelli determinati nel sistema bovino, dove era evidente una maggiore reattività del sistema misto di circa 6 volte, si rileva in fegato di ratto la pressochè identica reattività delle vie “mista” e “solubile”. La specificità delle attività “mista” e “solubile” nell’impiego della oloCRBPI per la sintesi di RA, viene evidenziata dal fatto che estratti freschi di cellule di lievito, che pure contengono elevati livelli di alcool e aldeide deidrogenasi, non sintetizzano RA da oloCRBPI o da retinaldeide generata da microsomi di bue o ratto. Seppure in via preliminare, sembra inoltre confermarsi l’effetto inibitorio della apoCRBPI, aggiunta in concentrazione equimolare alla miscela di reazione, sulla sintesi di acido retinoico anche nella frazione citosolica di ratto, oltre a essere evidente un sensibile calo nel sistema misto. L’impiego contemporaneo di apo e olo CRBPI quale substrato, oltre a minimizzare la quantità di retinolo in equilibrio, simula in modo appropriato le condizioni intracellulari, dove entrambe queste forme di CRBPI sono presenti.La quantità di acido retinoico sintetizzata nel sistema “misto” potrebbe in realtà includere anche la quota di RA prodotta dal sistema “solubile”, se si assume che il complesso retinolo-CRBPI possa essere convertito sia dalla deidrogenasi microsomale del sistema “misto”, che dall’attività del sistema “solubile” presente nella frazione citosolica impiegata. Questa possibilità verrebbe esclusa se l’inibizione osservata in presenza di minime quantità di proteine microsomali sulla sintesi citosolica di RA in ratto a partire da retinolo in forma libera riportata da Napoli e Race (Napoli e Race, 1990b) fosse presente anche nelle nostre condizioni di reazione, impiegando oloCRBPI come substrato.Come mostrato in Figura 17, l’aggiunta di quantità crescenti di microsomi ha effetti completamente differenti sulla formazione di RA nel sistema “solubile” di citosol di ratto e bue: nel primo l’inibizione della sintesi di RA determinata dalla presenza dei microsomi è estremamente marcata, e già dopo l’aggiunta di 20 g (1/10 della proteina citosolica), si osserva oltre l’80% di calo dell’attività; viceversa, tale effetto inibitorio è completamente assente in frazioni bovine, dove l’aggiunta si accompagna anzi ad un minimo aumento della sintesi di RA, presumibilmente a causa della nota presenza nei microsomi di attività retinolo e retinaldeide ossidasiche. Questi dati permettono di affermare che l’acido retinoico sintetizzato nella via “mista” di fegato di ratto sia prodotto esclusivamente da quel sistema biosintetico, che è attivo circa quanto quello “solubile”, diversamente da quanto avviene nel bue, dove la mancata inibizione microsomale, fa sì che la quantità di RA prodotto nel sistema “misto” comprenda anche una quota (minoritaria) prodotta dalla via “solubile”.
42

Risultati
L’analisi della dipendenza delle due vie di sintesi dai cofattori ha portato a risultati identici in bue e ratto nel caso del sistema “misto” (NADP in microsomi e NAD in citosol), mentre la via RA biosintetica “solubile” in ratto è strettamente dipendente dalla presenza di NAD (è attiva solo per il 10% in presenza di NADP), a differenza di quella in citosol di bue, che procede con una minore selettività nei confronti del coenzima (è circa doppia in presenza di NAD rispetto a quando è impiegato NADP) (Dati non mostrati).
Title: D:\CACCA\FIG05.PSCreator: GSS*CGI PostScript DriverCreationDate: 4 February 1996 17:54
Figura 17: Effetto dei microsomi sulla sintesi di acido retinoico nei sistemi “solubili” di bue e ratto. A 200 g di proteina citosolica di bue o ratto sono aggiunte quantità crescenti di microsomi omologhi. Le reazioni sono incubate in condizioni standard di reazione, con 4 M oloCRBP come substrato, e 2 mM NAD come cofattore.
Avendo osservato una minore reattività del sistema “misto” di ratto rispetto a quello di bue, abbiamo misurato la sintesi di RA da olo e apoCRBPI tentando la complementazione dell’attività di frazioni eterologhe, per identificare quale fosse, tra quella citosolica e microsomale, la deidrogenasi primariamente responsabile della differente velocità di reazione osservata.Come riportato in Tabella 8, la sintesi di RA nel sistema “misto” costituito da frazioni di specie diverse è possibile, e fornisce importanti elementi sulla velocità relativa delle attività enzimatiche microsomali e citosoliche: la retinaldeide deidrogenasi presente nel citosol di ratto, indipendentemente dalla specie dei microsomi con cui è complementata, è meno reattiva rispetto a quella bovina, mentre non sono rilevabili differenze significative tra l’attività retinolo deidrogenasica microsomale di frazioni da ratto e bue
43
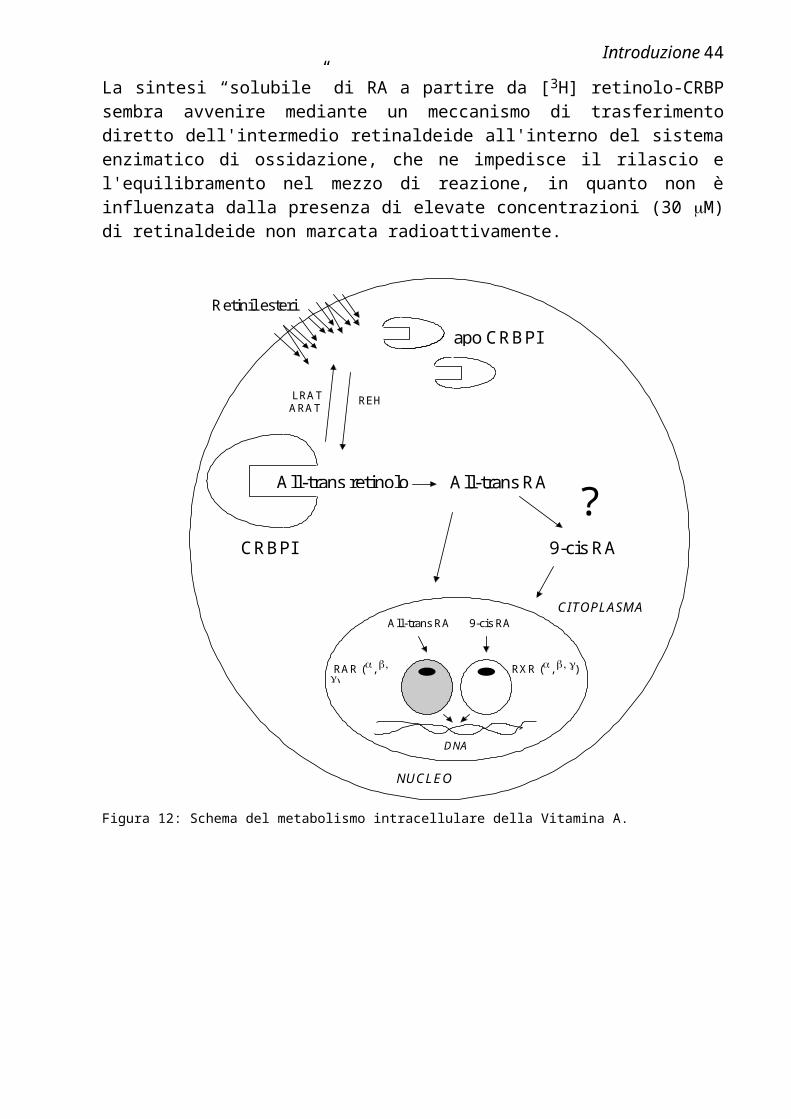
Risultati
sia quando sono complementate le RoDH delle due specie con lo stesso citosol, sia quando sono incubati i soli microsomi, ed è misurata la formazione di retinaldeide.
Frazione Acido retinoico
(pmoli)
Retinaldeide
(pmoli)
Microsomi Bue 4 68
Microsomi Ratto 5 53
Cit.+ Mic. Bue 184 10
Cit. + Mic. Ratto 12 27
Cit. Bue + Mic. Ratto 156 5
Cit. Ratto + Mic. Bue 37 18
Tabella 8: Sintesi di acido retinoico e retinaldeide in frazioni omologhe o eterologhe da fegato di bue o ratto. Le reazioni sono effettuate in condizioni standard di reazione (30’, 37°C, 2 mM NAD e/o NADP), impiegando 4 M olo e 4 M apoCRBPI con 200 g di proteina microsomale o 200 + 200 g di proteina microsomale e citosolica.
Questi dati indicano quindi che la minore reattività del sistema “misto” di ratto rispetto a quello bovino è presumibilmente causata dal componente solubile, la retinaldeide deidrogenasi, che determina la velocità di sintesi di RA quando complementata con microsomi omologhi o eterologhi, e non dalla retinolo deidrogenasi presente nella frazione microsomale, che non presenta differente attività in frazioni di ratto e bue.Un importante elemento che caratterizza la sintesi “solubile” in bue da oloCRBPI, e la differenzia da quella del sistema “misto”, è costituito dall’assenza di quantità rilevabili dell’intermedio di reazione retinaldeide. Abbiamo così verificato nel sistema “solubile” di ratto la presenza dell’intermedio, impiegando quale substrato anche retinolo in forma libera.
Specie SubstratoAcido retinoico
(pmoli)
Retinaldeide
(pmoli)
Bue oloCRBPI 30 N.D.
Bue Retinolo 230 2
Ratto oloCRBPI 34 6
Ratto Retinolo 155 41
Tabella 9: Sintesi citosolica di RA e RALD da retinolo in forma libera o legata a CRBPI. Le reazioni sono incubate per 30’ a 37°C, in presenza di 2 mM NAD come cofattore. La concentrazione di substrato è di 4 M oloCRBP e 7 M retinolo in forma libera, e la quantità di proteina citosolica impiegata è di 400 g.
44

Risultati
I valori ottenuti (Tabella 9) indicano la presenza nel citosol di ratto di quantità chiaramente rilevabili dell’intermedio di reazione retinaldeide, che sono sensibilmente maggiori quando è incubato retinolo in forma libera come substrato. A conferma dei dati precedentemente ottenuti impiegando [3H] retinolo-CRBP come substrato, l’accumulo di retinaldeide è minimo, o al di sotto dei limiti di rivelazione, nel caso del sistema “solubile” da fegato bovino.Oltre che dalla diversità dei due sistemi enzimatici “solubili” in specie diverse, il differente comportamento nell’accumulo dell’intermedio in citosol di ratto e di bue potrebbe essere semplicemente spiegato, nel caso che la via “solubile” fosse composta da due distinte attività enzimatiche, dalla differente velocità di ossidazione relativa del secondo componente, che risulterebbe essere quello limitante nel ratto, ed estremamente reattivo invece nel bue.Questi dati rivelano quindi nel loro complesso la presenza di entrambi i sistemi enzimatici, in fegato di bue e ratto capaci di sintetizzare RA a partire da oloCRBPI, sebbene con alcune differenze, minori, quali le diverse velocità relative, nella via “mista”, significative, invece, per la via “solubile”, dove la diversa specificità per il coenzima, così come l’accumulo o meno dell’intermedio di reazione costituiscono importanti caratteri distintivi.
4.5 Ipotesi sulla formazione dell’ acido 9-cis retinoico
La scoperta, avvenuta nel 1992, che l’acido 9-cis retinoico è il ligando naturale di RXR, ed è quindi coinvolto nel processo di attivazione trascrizionale di tutti i recettori nucleari eterodimerici, ha subito sollevato il problema della sua origine biologica.Il 9-cis RA è stato identificato quale componente endogeno in diversi tessuti adulti ed embrionali (fegato e reni di topo adulto; cervello e occhi di embrione di ratto), sebbene sia presente in concentrazioni da due a sei volte minori di quelle di all-trans RA, che attiva la trascrizione di distinti recettori nucleari (Heyman et al., 1992; CreechKraft e Juchau, 1993). Non è stata fino ad oggi ottenuta alcuna evidenza sperimentale che consenta di fornire una spiegazione plausibile riguardo la formazione di questo isomero dell’acido retinoico.Alcune ipotesi sono state fatte riguardo l’origine del 9-cis RA (Figura 18). Questa importante molecola potrebbe formarsi attraverso un processo di isomerizzazione post-sintetica di acido all-trans retinoico catalizzato da specifiche isomerasi, in modo analogo a quanto avviene nel sistema visivo, dove l’isomerizzazione dei retinoidi riveste un ruolo essenziale.Un’altra possibilità è che il 9-cis RA possa essere sintetizzato attraverso una reazione metabolica di ossidazione. Il complesso fisiologico all-trans retinolo-CRBP potrebbe fungere da substrato per una reazione di ossidazione e di contemporanea isomerizzazione
45

Risultati
da parte di uno dei due sistemi enzimatici, “solubile” o “misto” identificati in frazioni subcellulari di bue o ratto. In alternativa, è stata avanzata la possibilità che il 9-cis RA si possa formare mediante l’ossidazione di precursori aventi configurazione isomerica 9-cis: il 9-cis retinolo, la presenza del quale è stata di recente rilevata in reni di ratto (Labrecque et al., 1995), potrebbe essere un potenziale substrato per la sintesi di 9-cis RA.
ACIDO 9-CIS RETINOICO Acido all-trans retinoico
ISOMERIZZAZIONEPOST-SINTETICA
All-trans retinolo-CRBPI Precursori 9-cis (9-cis retinolo)
OSSIDAZIONE EISOMERIZZAZIONE
OSSIDAZIONE
Figura 18: Schema delle possibili vie di formazione del acido 9-cis retinoico.
4.6 Isomerizzazione di acido all-trans retinoico a acido 9-cis retinoico
Per verificare sperimentalmente la validità delle ipotesi sopra elencate, è stato necessario mettere a punto un sistema di analisi in HPLC che permettesse la separazione dei principali isomeri dell’acido retinoico. L’impiego di una nuova colonna C18 ad alta risoluzione, unitamente all’individuazione di una specifica fase mobile (Vedi Materiali e metodi), ci ha permesso di ottenere la separazione degli isomeri all-trans, 13-cis e 9-cis dell’acido retinoico a linea di base in poco più di 15 minuti (Figura 19).La prima ipotesi da noi verificata è stata la possibile formazione proteina-dipendente di 9-cis RA in seguito ad isomerizzazione di acido all-trans retinoico.A tale scopo, 200 pmoli di all-trans RA sono state incubate a 37°C per 60 minuti in presenza di frazioni citosoliche e microsomali di ratto (200 g di proteina ciascuna). La percentuale di isomero 9-cis formatasi in presenza delle frazioni subcellulari non è risultata significativamente maggiore di quella generata in seguito al solo processo di
46

Risultati
estrazione, e non è stata considerata quindi rappresentativa di una attività isomerasica (Tabella 10).
47
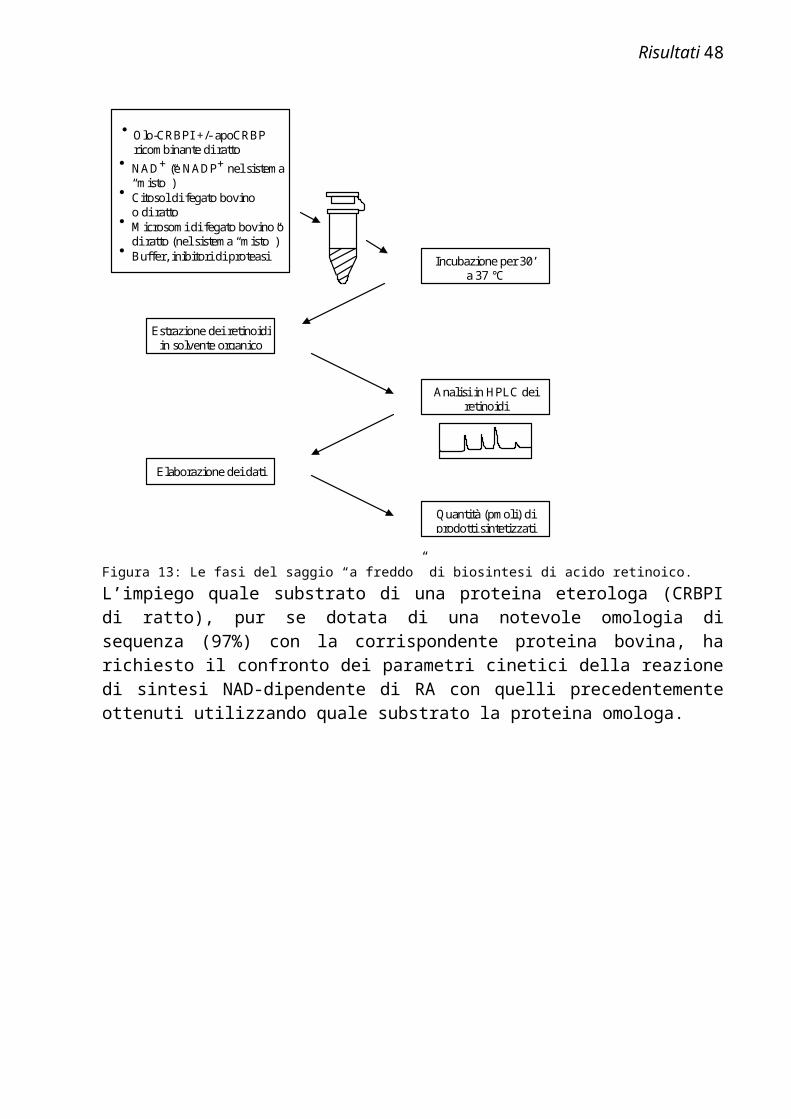
Risultati
Figura 19: Eluizione mediante HPLC di differenti isomeri di RA. Differenti quantità di standard all-trans, 13-cis e 9-cis RA sono stati separati impiegando la fase mobile “A”. I picchi eluiti corrispondono rispettivamente a 13, 9 e all-trans RA. La rilevazione è stata effettuata a 350 nm.
Estrazione IncubazioneAll-trans
(%)
13-cis
(%)
9-cis
(%)
- - 97 3 N.D.
+ - 70.6 27.1 2.1
+ + 69.2 27.9 2.9
Tabella 10: Isomerizzazione di acido all-trans retinoico. I dati riportati sono le medie di almeno 3 esperimenti eseguiti incubando 200 pmoli di acido all-trans retinoico, disperso in 1% etanolo, in presenza di 200 µg di proteine citosoliche e 200 µg di proteine microsomali da fegato di ratto più NAD e NADP (2 mM). E’ stata impiegata la procedura di estrazione con esano + etanolo/HCl (Vedi Materiali e metodi).
I medesimi risultati riguardo alla percentuale di 9-cis RA formatosi per isomerizzazione sono stati ottenuti nelle medesime condizioni di incubazione con proteine citosoliche e microsomali da fegato bovino. I dati da noi ottenuti non sono quantitativamente e qualitativamente dissimili da quelli riportati da Urbach e Rando, i quali rilevano la formazione di 9-cis RA incubando l’isomero all-trans dell’acido retinoico in presenza di microsomi bovini, o di sieroalbumina (1%) e detergenti (1% SDS) (Urbach e Rando, 1994a; Urbach e Rando, 1994b). Secondo questi autori tale processo può avere una qualche rilevanza fisiologica, ma le condizioni di reazione da essi impiegate sono lontane da quelle presenti in vivo, e, anche in questo caso, l’isomerizzazione di all-trans RA procede in modo proteina-indipendente. Poichè alcune proteine leganti composti idrofobici appartenenti alla famiglia della lipocaline possiedono attività catalitica di tipo isomerasico (Flower, 1994), è stata verificata la possibilità che la CRABPI potesse agire da catalizzatore per l’isomerizzazione di all-trans RA a 9-cis RA.
All trans RA Olo CRABPI Apo CRABPIAll-trans
(%)
13-cis
(%)
9-cis
(%)
+ - - 77 20 3
+ - + 78.2 18 3.8
- + - 79 18 3
- + + 73 23.3 3.6
Tabella 11: Isomerizzazione di acido all-trans retinoico in presenza di apoCRABPI. I dati riportati sono le medie di almeno 3 esperimenti eseguiti incubando 200 pmoli di acido all-trans retinoico, disperso in 1%
48

Risultati
etanolo, o legate a olo CRABP. Sono state impiegate le medesime condizioni di incubazione e la stessa procedura di estrazione indicata nella legenda della Tabella 10.
Come mostrato in tabella 11, l’analisi della configurazione isomerica di 200 pmoli di all-trans RA in forma libera o legato alla CRABPI, incubato per 60’ a 37°C con citosol e microsomi di ratto (200 g ciascuno), in presenza o in assenza di 10 M apoCRABPI, non ha portato ad un recupero significativamente differente di 9-cis RA in nessuno degli esperimenti effettuati.Sebbene sia possibile che l’attività acido retinoico isomerasica in presenza di frazioni citosoliche e microsomali, oltre che della CRABPI, possa procedere solo in presenza di effettori a noi ignoti, i dati a nostra disposizione, che indicano una pressoché nulla formazione di acido 9-cis retinoico, suggeriscono quindi che tale processo non sia fisiologicamente importante per la biosintesi di acido 9-cis retinoico.
4.7 Analisi della configurazione isomerica dell’acido retinoico sintetizzato in presenza dei sistemi “misto” e “solubile” a partire da all-trans retinolo-CRBPI
Un’altra possibile via di formazione di 9-cis RA potrebbe essere costituita da una reazione enzimatica di contemporanea ossidazione e isomerizzazione che impiega come substrato il complesso fisiologico all-trans retinolo-CRBPI. Esistono esempi noti di enzimi, quale ad esempio la 3-idrosteroide deidrogenasi/ 4-5 isomerasi NAD-dipendente, coinvolta nella biosintesi di tutte le classi di steroidi attivi biologicamente, in grado di catalizzare una reazione di ossidazione e isomerizzazione (Rheaume et al., 1995). E’ stata quindi analizzata la forma isomerica dell’acido retinoico sintetizzato in presenza dei sistemi “misto” e “solubile”. Infatti, l’eventuale capacità di ognuna della due vie di sintetizzare prodotti aventi distinta configurazione isomerica (all-trans e 9-cis RA), potrebbe fornire una plausibile spiegazione dell’esistenza di due distinte attività acido retinoico biosintetiche che utilizzano il medesimo substrato, oloCRBPI.Risultati preliminari da noi ottenuti con frazioni di ratto e bue sembravano mostrare una distinta stereoselettività dei due sistemi biosintetici (Cavazzini et al., 1994). Utilizzando quale substrato 4 M all-trans retinolo-CRBP, in presenza di uguali concentrazioni di apo CRBP al fine di mimare condizioni di reazione intracellulari, il sistema "misto" manteneva la configurazione all-trans a livello del prodotto, mentre l'attività della frazione solubile era in grado di generare selettivamente l'isomero 9-cis dell'acido retinoico (Figura 20).Il risultato ottenuto era estremamente interessante, perchè evidenziava una possibile via di sintesi dell’acido 9-cis retinoico a partire da oloCRBPI in condizioni “fisiologiche”, e consentiva di fornire una plausibile spiegazione della presenza di due distinti sistemi ossidativi.
49

Risultati
Numerosi controlli sono stati eseguiti per verificare la validità dei dati ottenuti, tra cui un'accurata analisi mediante HPLC della configurazione stereoisomerica dell'all-trans retinolo contenuto in diversi pool di oloCRBP utilizzata per le reazioni biosintetiche
oloCRBPI +apoCRBPI
All-trans RASistema“misto”
9-cis RASistema“solubile”
Figura 20: Forma isomerica dell’acido retinoico sintetizzato nel sistema “misto” e “solubile” di fegato di ratto.
in vitro, che ha evidenziato la presenza di una piccola, ma significativa percentuale (3%) di retinolo 9-cis, formatosi durante il processo di purificazione e risaturazione della proteina ricombinante. Nonostante fosse presente in quantità assai modeste, il 9-cis retinolo preesistente, se reattivo, avrebbe potuto influenzare in modo determinante la apparente specificità stereoisomerica dei due sistemi ossidativi.Sono così state apportate alcune modifiche mirate alla procedura di formazione del complesso retinolo-CRBPI, che ci hanno permesso di preparare CRBP contenente esclusivamente all-trans retinolo. In questo caso il ligando, la cui purezza era stata verificata mediante analisi in HPLC, veniva aggiunto solo prima dell'ultima fase di purificazione della CRBPI di ratto, minimizzando così eventuali processi di isomerizzazione (Vedi Materiali e metodi).
4.8 Il complesso 9-cis retinolo-CRBPI
La presenza di 9-cis retinolo in vari pool di CRBPI, sebbene quantitativamente minoritaria, era difficilmente giustificabile sulla base dei dati presenti in letteratura. Alcuni autori avevano infatti riportato l'assoluta selettività di legame della CRBPI di ratto per l'isomero all-trans del retinolo, e l’apparente mancato riconoscimento del ligando in forma 9-cis (MacDonald e Ong, 1987).D’altra parte, l’impiego nel sistema “solubile” di ratto di apo e oloCRBPI che erano state sottoposte ad un’ulteriore cromatografia per filtrazione molecolare in grado di rimuovere
50

Risultati
eventuali retinoidi liberi presenti anche in tracce, non ha evidenziato alcuna differenza nella sintesi di 9-cis RA rispetto a quando era utilizzata oloCRBPI non trattata.Analogamente, la formazione di 9-cis RA nel sistema “solubile” viene abolita (si recupera esclusivamente all-trans RA) impiegando quale substrato oloCRBPI che era stata incubata per 3 ore con un eccesso 10 molare di all-trans retinolo, seguita dalla rimozione del ligando in eccesso, in presenza di una quantità equimolare di apoCRBPI.Nel loro insieme queste evidenze ci hanno portato ad ipotizzare che la formazione di acido 9-cis retinoico fosse causata dalla conversione di 9-cis retinolo legato (o adsorbito) alla CRBPI, ma non presente in equilibrio in forma libera. Avendo a nostra disposizione numerose indicazioni a favore dell’esistenza di un’interazione specifica tra 9-cis retinolo e CRBPI, abbiamo quindi tentato l’isolamento del complesso.Lo spettro di assorbimento UV-visibile della CRBPI risaturata con 9-cis retinolo (preparato per riduzione della 9-cis retinaldeide), e sottoposta a gel-filtrazione per rimuovere l’eccesso di ligando, presenta il tipico profilo di una proteina in forma olo (Figura 21), con caratteristiche spettrali del ligando riconducibili al 9-cis retinolo.
51

Risultati
Figura 21: Spettri di assorbimento UV-visibile del complesso 9-cis retinolo-CRBPI ( ¾ ) e all-trans retinolo-CRBPI ( - - - -).
Lo spettro di assorbimento del complesso 9-cis retinolo-CRBPI presenta un massimo a 332 nm, ed ha un andamento caratteristico, distinto da quello del complesso all-trans retinolo-CRBPI. Tale complesso risulta tuttavia piuttosto instabile in soluzione, a causa di una lenta conversione del ligando dalla forma 9-cis ad all-trans (t1/2 @ 14 giorni a 4°C), rilevabile sia dall’analisi dello spettro di assorbimento che mediante HPLC. Quando impiegato per prove di ossidazione in vitro, il complesso 9-cis retinolo-CRBPI è stato quindi utilizzato immediatamente dopo la sua preparazione.E’stata anche misurata, mediante misure di quenching della fluorescenza dei triptofani della CRBPI in seguito al legame con il retinoide, l’interazione tra proteina legante e 9-cis retinolo (Figura 22). La costante di dissociazione stimata (circa 10-8 M), anche se è circa 100 volte maggiore di quella misurata nel caso dell'all-trans retinolo, è indice di una buona affinità della proteina per il 9-cis retinolo, e conferma in modo diretto l’esistenza di uno specifico legame tra questo composto e la CRBPI.Il differente comportamento osservato rispetto ai dati riportati in letteratura è probabilmente attribuibile al diverso metodo di analisi impiegato. Mac Donald e Ong, infatti, hanno misurato il legame di vari retinoidi con la CRBPI, tra cui il 9-cis retinolo,
Title: D:\CACCA\FIG07.PSCreator: GSS*CGI PostScript DriverCreationDate: 4 February 1996 17:57
Figura 22: Curva di legame del 9-cis retinolo alla CRBPI. L’analisi è stata eseguita misurando il “quenching” di fluorescenza della proteina a l280 ad una concentrazione di 1µM CRBPI.
determinando l’aumento dell’intensità di fluorescenza in seguito alla formazione del complesso. Tale metodo non è tuttavia applicabile all’isomero 9-cis del retinolo
52

Risultati
poichè abbiamo osservato che questo, a differenza di altri isomeri tra i quali l’all-trans retinolo, non aumenta la propria fluorescenza a seguito del legame con la proteina.4.9 Stereoselettività delle vie di sintesi “mista” e “solubile” nella formazione di RA
da all-trans retinolo in forma libera o legata a CRBPI
Avendo identificato all’interno dei pool di oloCRBPI precedentemente impiegati la presenza di due distinte forme isomeriche di retinolo (all trans e 9-cis, entrambe legate alla proteina), che impediscono di ottenere un’interpretazione chiara ed univoca dei risultati ottenuti riguardo alla stereospecificità dei sistemi enzimatici di ossidazione, si è reso necessario ripetere l’analisi della forma isomerica dell’acido retinoico prodotto in vitro nel sistema “solubile” e “misto“ di ratto e bue, impiegando complessi proteici contenenti esclusivamente all-trans o 9-cis retinolo.Sebbene i retinoidi in forma legata alle binding proteins costituiscano il probabile substrato in vivo per le diverse reazioni in cui è coinvolta la vitamina A, tutte le evidenze sperimentali fino ad oggi riportate indicano che i diversi sistemi enzimatici, anche in forma altamente purificata, che riconoscono il complesso retinolo-CRBPI, risultano comunque sempre in grado di metabolizzare i retinoidi in forma libera. Abbiamo pertanto impiegato all-trans e 9-cis retinolo dispersi in solvente organico per valutare la stereoselettività dell’attività retinoico biosintetica delle vie “solubile” e “mista” in frazioni subcellulari da fegato di ratto e bue, e confrontare i risultati così ottenuti con quelli raccolti in presenza del complesso retinolo-CRBPI. E’ stata saggiata inizialmente la reattività del complesso CRBPI all-trans retinolo, e la configurazione isomerica dell’acido retinoico prodotto a partire da tale substrato in presenza di frazioni subcellulari isolate da fegato di ratto. A conferma di quanto osservato in precedenza (Tabella 7), la formazione di acido retinoico in presenza di concentrazioni fisiologiche di oloCRBPI è rilevabile in entrambi i sistemi, “solubile” e “misto”, che mostrano simile reattività (Tabella 12).
Apo-CRBPISistema
enzimaticoCofattore
Acido Retinoico (pmoli)
Configurazione isomerica
- Solubile NAD 25 All-trans
- Solubile NADP 4.8 All-trans
- Misto NAD + NADP 20 All-trans
+ Solubile NAD 1.6 All-trans
+ Misto NAD + NADP 11 All-trans
Tabella 12: Sintesi di acido retinoico da all-trans retinolo-CRBPI in frazioni subcellulari da fegato di ratto. Le reazioni sono state eseguite incubando in condizioni standard di reazione 4 µM olo-CRBPI e, quando presente, 6 µM di apoCRBPI, impiegando 400 µg di proteine citosoliche per il sistema “solubile” o 200 +
53

Risultati
200 µg di proteine citosoliche e microsomali per il sistema “misto”. La concentrazione dei cofattori è di 2 mM.
Viene inoltre confermata la maggiore selettività per il coenzima dell’attività “solubile” di ratto rispetto a quella di bue: meno del 20% dell’acido retinoico viene prodotto in citosol di ratto in presenza di NADP, rispetto a quando è utilizzato NAD come cofattore.In queste nuove condizioni, l’analisi della configurazione isomerica del prodotto di reazione ha evidenziato la formazione della sola forma all-trans dell’acido retinoico in entrambe le vie biosintetiche da frazioni di fegato di ratto, escludendo l’esistenza un possibile processo di contemporanea ossidazione ed isomerizzazione dell’all-trans retinolo legato alla CRBPI. La medesima forma isomerica di acido retinoico è stata rilevata anche quando era incubato come substrato all-trans retinolo-CRBPI in presenza di un eccesso di apoCRBPI per meglio simulare condizioni intracellulari (Tabella 12). Sorprendentemente, l’effetto inibitorio della apoCRBPI è molto maggiore di quello osservato precedentemente nel caso del sistema “solubile” (cfr. Tabella 7 e 12). In condizioni di reazione in cui la sintesi di RA da parte del sistema “misto” è inibita (ca. 50%), ma chiaramente rilevabile, nel sistema “solubile” da fegato di ratto, la sintesi di RA risulta di poco superiore ai limiti di rivelazione (ca. 95% di inibizione apparente). Questi risultati, che evidenziano come l’ossidazione di all-trans retinolo legato a CRBPI produca acido retinoico avente la medesima configurazione isomerica del substrato, sono stati confermati impiegando all-trans retinolo in forma libera quale substrato. Anche in questo caso infatti, oltre alla sintesi selettiva di all-trans RA, si è verificato l’impiego preferenziale del NAD come cofattore da parte della via biosintetica “solubile” (Tabella 13).
Sistemaenzimatico
CofattoreAcido Retinoico
(pmoli)Configurazione isomerica
Solubile NAD 60 All-trans
Solubile NADP 8.2 All-trans
Misto NAD + NADP 26.9 All-trans
Tabella 13: Sintesi di acido retinoico da all-trans retinolo in forma libera in frazioni subcellulari da fegato di ratto. Le reazioni sono state eseguite incubando 5 µM di retinolo libero solubilizzato in 1% di etanolo in condizioni standard di reazione, in presenza di 400 µg di proteine citosoliche per il sistema “solubile” o 200 + 200 µg di proteine citosoliche e microsomali per il sistema “misto”. La concentrazione dei cofattori è di 2 mM.
Anche impiegando frazioni citosoliche e microsomali da fegato bovino, l’acido all-trans retinoico è l’unico prodotto di reazione sia a partire da all-trans retinolo disperso in solvente organico, sia in forma legata alla CRBPI. Anche in questo caso, l’inibizione
54

Risultati
della sintesi di RA da parte della apoCRBPI è pressochè completa. L’unica differenza riguarda la minore selettività del sistema “solubile” nell’impiego del coenzima: sebbene il NAD sia il cofattore preferito, si osserva una sintesi consistente di RA anche in presenza di NADP.
4.10 Effetto della apoCRBPI sulla sintesi di RA da all-trans retinolo-CRBPI
Impiegando il nuovo preparato di all-trans retinolo-CRBPI, si è rilevato, nel caso del sistema “solubile” da fegato di ratto, un notevole effetto inibitorio della apoCRBPI sulla sintesi di RA, più consistente di quello rilevato in precedenza impiegando preparati di oloCRBPI microeterogenei (contenenti 3-5% di 9-cis retinolo) come substrato. Questo differente comportamento ci ha indotto ad analizzare in maggior dettaglio l’effetto delle apo proteine sulla formazione di acido retinoico nei due sistemi biosintetici.Come indicato dalle curve di inibizione riportate in Figura 23, l’aggiunta di quantità crescenti di apoCRBPI in presenza di una concentrazione costante di all-trans retinolo-CRBPI (2 M), determina una netta diminuzione della sintesi di RA in entrambi i sistemi.
Title: D:\CACCA\FIG08.PSCreator: GSS*CGI PostScript DriverCreationDate: 4 February 1996 18:00
Figura 23: Effetto della apoCRBPI sulla sintesi di RA nei sistemi “solubile“ e “misto” di fegato di ratto. All-trans retinolo-CRBPI alla concentrazione di 2 M è incubata in condizioni standard di reazione in presenza di quantità crescenti di apoCRBPI. La quantità di proteina impiegata è di 400 g per il sistema “solubile” ( n ), e 200 + 200 g di microsomi e citosol nel caso si testi il sistema “misto” ( l ). La curva tratteggiata con il simbolo ( ¨ ) rappresenta la concentrazione stimata di retinolo in equilibrio con la oloCRBPI alle varie concentrazioni di apoCRBPI (100% retinolo libero: 44 nM); la curva tratteggiata con il simbolo ( ¡ ) è la curva attesa nel caso di una inibizione competitiva della apoCRBPI. La quantità di
55

Risultati
acido retinoico sintetizzato in assenza di apoCRBPI è pari a 10 e 12 pmoli nel caso del sistema “misto” e “solubile” rispettivamente.
Sebbene l’effetto inibitorio della apoCRBP nei confronti del sistema “solubile” sia nettamente più marcato di quanto osservato nel caso del sistema misto, i dati a disposizione non permettono di distinguere con certezza se tale fenomeno sia causato dal sequestramento di retinolo in equilibrio (da parte della apoCRBPI) (Napoli et al., 1991),oppure da una inibizione competitiva della apo sulla olo CRBPI rispetto ad un numero limitato di siti enzimatici (Ottonello et al., 1993). La curva di inibizione teorica1 costruita ipotizzando che il retinolo libero in equilibrio, piuttosto che la oloCRBP, rappresenti il vero substrato di ossidazione (assumendo Kdiss= 10-10 M), risulta quasi sovrapponibile
alla curva sperimentale osservata nel caso del sistema “solubile”, mentre la curva di inibizione calcolata assumendo l’esistenza di una inibizione competitiva tra apo e olo CRBPI (assumendo Ki= Km) ha un andamento similare2 , anche se non perfettamente
sovrapponibile, a quella ottenuta sperimentalmente nel caso del sistema “misto”. Nonostante ciò, diverse considerazioni consigliano prudenza nell’affermare che queste due ipotesi siano quelle valide per spiegare l’inibizione da apoCRBP dei due sistemi biosintetici. L’impiego di valori di Kdiss e Ki leggermente diversi da quelli assunti per
calcolare le curve di inibizione, ha come conseguenza l’ottenimento di curve teoriche notevolmente diverse. Inoltre, l’ipotesi che il retinolo in equilibrio sia l’effettivo substrato del sistema enzimatico “solubile” è in contrasto con il fatto che la dipendenza della velocità di sintesi dalla concentrazione di oloCRBP è di tipo saturabile, con un valore di Km (0.84 M), precedentemente determinato, compatibile con la concentrazione
di oloCRBPI presente in diversi tessuti. Quest’ultimo dato, assieme all’osservazione che le concentrazioni di retinolo in equilibrio stimate sulla base dell’equazione (1) sono di almeno due ordini di grandezza inferiori alla Km determinata per il retinolo libero (7
1
1La curva teorica di inibizione del retinolo in equilibrio è stata ricavata impiegando la seguente equazione:
RlR t Kdiss
Kdiss CRBP t R t
dove Rl è la concentrazione di retinolo in equilibrio, [R]t la concentrazione di retinolo totale, Kdiss la costante di dissociazione, e [CRBP]t la concentrazione totale di CRBPI.
2 La velocità di sintesi di RA nel sistema “solubile e “misto” in presenza di un inibitore competitivo è stata calcolata impiegando la seguente equazione:
V
V Ki S
Ki S Km Ki Km Imax
dove Vmax e Km sono i parametri cinetici della reazione, Ki rappresenta la costante di inibizione, mentre [ I ] è la concentrazione di inibtore.
56

Risultati
µM), indicherebbe piuttosto l’esistenza di una qualche forma di riconoscimento specifico tra CRBPI ed enzima ossidante “solubile”.Pertanto, sebbene non sia chiaro il meccanismo molecolare che sta alla base della azione inibitoria della apoCRBP, risulta evidente (almeno a livello empirico) che il sistema “misto” è meno suscettibile del sistema solubile ad inibizione da apo-proteina ed è quindi in grado di funzionare, sia pure a livelli sub-massimali, anche in presenza di concentrazioni relativamente elevate di apoCRBP. Al contrario, il sistema “solubile” è in grado di formare RA quasi unicamente in presenza del complesso stechiometrico all trans retinolo-CRBPI.Poichè i risultati dell’analisi strutturale del complesso retinolo-CRBP indicano che il retinolo deve in qualche modo essere rilasciato dal -barrel per poter essere reattivo (il retinolo in forma libera viene convertito in retinaldeide anche dalla retinolo deidrogenasi microsomale in forma altamente purificata), e dal momento che questo evento sembra costituire lo stadio limitante dell’intero processo di ossidazione ad acido retinoico, l’inibizione da apoCRBP potrebbe in realtà riflettere una competizione fra sito attivo dell’enzima e apoCRBP (in eccesso) per il retinolo rilasciato dalla olo-proteina a seguito dell’interazione con la deidrogenasi solubile o microsomale. Pertanto, una diversa affinità dei due enzimi per il retinolo rilasciato, e la loro differente concentrazione locale, oltre al fatto che la membrana microsomale potrebbe agire da sink aspecifico per la apoCRBP (che è nota avere una elevata affinità per le membrane) - riducendo così la micro-concentrazione di apoCRBP in prossimità del sito attivo - potrebbero tutte contribuire a spiegare la diversa risposta dei due sistemi di ossidazione ad un uguale eccesso di apoCRBP. Seppure estremamente plausibile, quest’ultimo modello non può al momento essere verificato dal punto di vista sperimentale poichè non si conoscono le affinità relative dei due enzimi per il retinolo libero. Inoltre, la mancanza di anticorpi diretti contro la deidrogenasi solubile impedisce di ottenere una stima attendibile della concentrazione dei due enzimi nelle rispettive frazioni subcellulari da fegato di ratto.
4.11 Stereoselettività delle vie di sintesi “mista” e “solubile” per la formazione di RA da 9-cis retinolo in forma libera o legata a CRBPI
Avendo escluso la possibilità che 9-cis RA si formi per isomerizzazione post-sintetica, o attraverso un contemporaneo processo di ossidazione ed isomerizzazione, abbiamo successivamente verificato la possibilità che la formazione di tale isomero dell’acido retinoico possa procedere attraverso l’ossidazione di 9-cis retinolo.Il 9-cis carotene, un componente minore della dieta, assorbito a livello intestinale, costituisce il precursore del 9-cis retinolo in fegato di ratto e pollo (Ben-Anotz et al., 1988; Levin e Mokady, 1994). Sebbene non si conoscano nè le modalità nè i siti preferenziali di tale conversione, vari studi hanno dimostrato che il 9-cis retinolo è un
57

Risultati
componente endogeno di diversi organi di ratto (reni, fegato) (Labrecque et al., 1995; Ben-Anotz et al., 1988), e umani (fegato, reni, testicoli) (Stahl et al., 1993).La reattività del 9-cis retinolo, legato alla CRBPI o disperso in solvente organico, unitamente alla configurazione isomerica dell’acido retinoico eventualmente sintetizzato attraverso le vie biosintetiche “solubile” e “mista”, sono state studiate in frazioni citosoliche e microsomali da fegato di ratto.Dopo incubazione di 9-cis retinolo-CRBPI 2 M (Tabella 14) in presenza di estratti solubili da fegato di ratto, si osserva sintesi di acido retinoico avente esclusivamente la configurazione isomerica 9-cis. Analogamente a quanto osservato utilizzando all-trans retinolo come substrato, questa attività enzimatica mostra una netta preferenza per il NAD come cofattore, rispetto al NADP (6% attività residua).
Sistema enzimatico CofattoreAcido retinoico
(pmoli)Configurazione
isomerica
Solubile NAD 48 9-cis
Solubile NADP 3 9-cis
Misto NAD + NADP 3 9-cis
Tabella 14: Sintesi di acido retinoico a partire da 9-cis retinolo CRBPI nel sistema “misto” e “solubile” di fegato di ratto. Le reazioni sono state eseguite in condizioni standard ( 2mM NAD e/o NADP) incubando 2µM di retinolo 9-cis retinolo-CRBP in presenza di 200 µg di proteina citosolica nel caso del sistema “solubile”, e 200 µg di proteina citosolica più 200 µg di proteina microsomale nel caso del sistema “misto”.
Il dato più sorprendente (Tabella 14), è la pressochè nulla reattività del complesso 9-cis retinolo-CRBPI nel sistema “misto” da fegato di ratto (Cfr. Tabella 14 e 12). I risultati ottenuti impiegando quale substrato 9-cis retinolo in forma libera (5 M) sono sostanzialmente identici a quelli ottenuti in presenza di CRBPI: il sistema “solubile” è
Sistema enzimatico CofattoreAcido retinoico
(pmoli)Configurazione
isomerica
Solubile NAD 100 9-cis
Solubile NADP 5 9-cis
Misto NAD + NADP 4.8 9-cis
aSolubile NAD 40 9-cis
Tabella 15: Sintesi di acido 9-cis retinoico da retinolo libero nel sistema “misto” e “solubile” di fegato di ratto. Le reazioni sono state eseguite incubando 5µM di 9-cis retinolo, in presenza di 200µg di solo
58

Risultati
citosol o di 200µg citosol + 200µg microsomi per il sistema misto in condizioni standard di reazione; la concentrazione dei cofattori è di 2 mM.
aIn questo caso il substrato, 5 M 9-cis retinolo, è stato incubato
in presenza di una concentrazione equimolare di apoCRBPI.
l’unico in grado di sintetizzare acido 9-cis retinoico a partire da 9-cis retinolo, ed impiega selettivamente il NAD come cofattore (Tabella 15). Come mostrato in Tabella 15, l’aggiunta di concentrazioni equimolari di apoCRBPI ad una miscela di reazione contenente 5 M retinolo disperso in 1% etanolo causa una inibizione di circa il 60% nella sintesi di 9-cis RA nel sistema solubile. Questo dato rappresenta un’ulteriore conferma dell’esistenza di uno specifico legame tra CRBPI e 9-cis retinolo, poichè, diversamente, non si sarebbe dovuto osservare alcun effetto inibitorio sulla sintesi di RA. Viene inoltre evidenziata la minore velocità di ossidazione del 9-cis retinolo quando è legato alla CRBPI, rispetto a quando è presente nella miscela di reazione in forma libera. Questo tipo di comportamento è comune a diversi tipi di reazioni nelle quali è coinvolto il complesso retinolo-CRBPI, ed è spiegabile dal fatto che il rilascio del retinolo dalla proteina è considerato il passaggio limitante dell’intero processo di conversione.Analoghi esperimenti effettuati con frazioni subcellulari da fegato bovino, hanno pure evidenziato una chiara conversione preferenziale dei substrati 9-cis e all-trans da parte dei sistemi “solubile“ e “misto”, con l’esclusiva formazione nel primo caso di acido 9-cis retinoico.Poichè l’attività biosintetica del sistema “misto” in frazioni isolate da fegato di ratto è ai limiti di rilevazione quando si impiega come substrato 9-cis retinolo in forma libera o associato a CRBPI, ci siamo chiesti quale dei due componenti, citosolico o microsomale, fosse responsabile della scarsa reattività osservata.I dati riportati in Tabella 16 evidenziano la pressochè totale incapacità dell’attività retinolo deidrogenasica microsomale da fegato di ratto di ossidare 9-cis retinolo nelle stesse condizioni in cui l’isomero all-trans viene efficacemente convertito in retinaldeide.
Substrato CofattoreRetinaldeide
(pmoli)Configurazione
isomerica
All-trans retinolo NAD 80 All-trans
All-trans retinolo NADP 118 All-trans
All-trans retinolo NAD + NADP 101 All-trans
9-cis retinolo NAD 7 9-cis
9-cis retinolo NADP 6 9-cis
9-cis retinolo NAD + NADP 7 9-cis
Tabella 16: Sintesi di retinaldeide in microsomi di fegato di ratto. 7 M di all-trans o 9-cis retinolo sono stati incubati in condizioni standard di reazione in presenza di 200 µg di proteina microsomale da fegato
59

Risultati
di ratto. Ai valori indicato è stata sottratta quantità di retinaldeide (7 pmoli) sintetizzata incubando i microsomi in presenza del solo cofattore.
L’analisi della selettività riguardo ai cofattori NAD e NADP, sebbene confermi l’impiego leggermente preferenziale del NADP in presenza di all-trans retinolo, non evidenzia alcun incremento coenzima-dipendente della sintesi di 9-cis retinaldeide a partire da 9-cis retinolo.Il medesimo esperimento, eseguito in presenza della frazione microsomale isolata da fegato bovino, ha confermato l’esistenza di un riconoscimento selettivo del retinolo all-trans (220 pmoli di all-trans retinaldeide prodotte con 2 mM NADP), e una minima reattività nei confronti del retinolo in forma 9-cis. Confrontando questi dati, con quelli recentemente riportati da El Akawi e Napoli (El Akawi e Napoli, 1994), riguardo alla specificità di substrato del secondo componente del sistema “misto” (retinaldeide deidrogenasi), che può efficacemente utilizzare quale substrato sia all-trans che 9-cis retinaldeide, risulta evidente come la selettività per l’isomero all-trans del sistema “misto” sia determinata esclusivamente dalla incapacità della retinolo-deidrogenasi microsomale di convertire il 9-cis retinolo.I risultati da noi ottenuti, evidenziano quindi, per la prima volta, l’esistenza di una via enzimatica di sintesi di acido 9-cis retinoico in frazioni subcellulari da fegato di ratto (e bue). Il sistema biosintetico “solubile” è l’unico in grado di utilizzare efficacemente il 9-cis retinolo, in forma libera o legata a CRBPI, quale substrato per la sintesi di 9-cis RA.Abbiamo effettuato alcuni esperimenti per poter caratterizzare l’attività 9-cis RA sintetasica presente nella frazione solubile da fegato di ratto, definire l’effetto della apoCRBPI, e ottenere maggiori informazioni sulla reattività del complesso 9-cis retinolo-CRBPI.L’aggiunta di concentrazioni crescenti di apoCRBPI in presenza di una concentrazione costante (1 M) del complesso 9-cis retinolo-CRBPI, provoca una chiara diminuzione della sintesi di acido 9-cis retinoico (Figura 24).Anche in questo caso, così come per l’analisi dell’inibizione da apoCRBPI sulla produzione di all-trans RA, per verificare se il calo della sintesi di 9-cis RA fosse causato dal sequestramento del retinolo in equilibrio, o dall’inibizione competitiva della apo sulla oloCRBP, sono state calcolate entrambe le curve teoriche di inibizione, assumendo un valore di Kdiss per il complesso 9-cis retinolo CRBPI pari a 1x10-8 M, e la costante di dissociazione enzima-inibitore Ki uguale a Km.
I dati sperimentali ottenuti, relativi all’inibizione della apoCRBPI sulla sintesi di 9-cis RA in presenza di 9-cis retinolo-CRBPI, non correlano con alcuna delle curve teoriche calcolate, sebbene quella relativa all’inibizione competitiva tra apo e oloCRBPI sia quella che maggiormente si avvicina ai valori osservati (Figura 24).L’analisi dei dati, pur senza chiarire quale sia il meccanismo di inibizione della apoCRBPI sulla sintesi del 9-cis RA, sembra escludere che la catalisi avvenga sul
60

Risultati
retinolo presente in equilibrio, anche se, per ottenere un’ulteriore conferma in questo senso, sono necessarie altre evidenze sperimentali, prima fra tutte una stima più accurata della
Title: D:\CACCA\FIG09.PSCreator: GSS*CGI PostScript DriverCreationDate: 4 February 1996 18:03
Figura 24: Effetto della apoCRBPI sulla ossidazione nel sistema “solubile” del complesso 9-cis retinolo-CRBPI. Le reazioni sono incubate in condizioni standard in presenza di una quantità costante di 9-cis retinolo-CRBPI (1 M) e di proteina citosolica (400 g), e concentrazioni crescenti di apoCRBPI.La curva tratteggiata con il simbolo ( ) rappresenta la concentrazione teorica di retinolo in equilibrio con la oloCRBPI in presenza di differenti concentrazioni di apoCRBPI (100% retinolo libero: 100 nM); la curva tratteggiata con il simbolo ( o ) indica l’inibizione attesa nel caso di una inibizione competitiva tra apo e olo CRBPI. La quantità di acido retinoico sintetizzato in assenza di apoCRBPI è di 47 pmoli.
costante di dissociazione tra il 9-cis retinolo e la CRBPI, il cui valore è essenziale nel calcolo della curva teorica di inibizione. E’ stata inoltre determinata la dipendenza della velocità di sintesi di 9-cis RA in presenza di concentrazioni crescenti di apo e olo CRBPI legante 9-cis retinolo, impiegate ad un rapporto costante di 3:1 (Figura 25).Sebbene la produzione di acido retinoico in queste condizioni sperimentali sia minore di quella misurabile in assenza di apoCRBPI, e il valore della Km apparente risulti
maggiore, la quantità di retinolo presente in equilibrio è praticamente costante per ogni valore di concentrazione della olo proteina.Ciò permette di distinguere se la formazione di acido retinoico dipende dal retinolo libero in equilibrio, nel qual caso la quantità di acido retinoico prodotto sarebbe costante, oppure dalla concentrazione del complesso retinolo-CRBP, nel qual caso la velocità di sintesi aumenterebbe all’aumentare della concentrazione della oloCRBPI.Come si può osservare in Figura 25, quest’ultimo caso è quello verificato sperimentalmente, in quanto si osserva un chiaro andamento saturabile della sintesi di
61

Risultati
acido 9-cis retinoico; i valori dei parametri cinetici apparenti determinati sono di 0.27 M per la Km, mentre la Vmax è di circa 135 pmoli/ h mg di proteina citosolica.
I dati relativi all’effetto dell’apoCRBP, unitamente alla curva di saturazione, suggeriscono quindi l’esistenza di un possibile riconoscimento preferenziale del
Title: D:\CACCA\FIG10.PSCreator: GSS*CGI PostScript DriverCreationDate: 4 February 1996 18:06
Figura 25: Dipendenza della sintesi di 9-cis RA dalla concentrazione di 9-cis retinolo-CRBPI. Olo e apo CRBPI ad un rapporto costante di concentrazione pari a 1:3 sono state incubate in condizioni standard di reazione in presenza di 400 g di proteina citosolica e 2 mM NAD come cofattore.
complesso 9-cis retinolo-CRBPI piuttosto che del retinolo in forma libera (sebbene ulteriori conferme sperimentali siano necessarie). I valori dei parametri cinetici (Km e Vmax) sono simili a quelli precedentemente riportati per reazioni che coinvolgono all-
trans retinolo-CRBP (ossidazione, esterificazione), ma nel caso del complesso 9-cis retinolo-CRBPI, per la prima volta isolato in vitro, non esiste alcun riferimento sulla sua presenza (e quindi sulla sua concentrazione) in vivo. Ciò impedisce qualsiasi considerazione sulla possibile rilevanza in vivo di tale reazione biosintetica, e sul possibile ruolo fisiologico del complesso. Si può semplicemente osservare che i dati riportati in letteratura indicano una concentrazione endogena di 9-cis retinolo che è circa dieci volte inferiore a quella dell’all-trans retinolo (circa 10-7 M in rene di ratto; Labrecque et al., 1995), ma non sono disponibili informazioni riguardo alla velocità con la quale esso viene metabolizzato nei tessuti.L’isolamento e la successiva caratterizzazione dell’attività solubile in grado di sintetizzare 9-cis RA, oltre a nuovi esperimenti volti a chiarire se il 9-cis retinolo in forma libera o legata alla CRBPI possa costituire un valido precursore in vivo, potranno
62

Risultati
chiarire l’effettiva importanza fisiologica della via biosintetica “solubile” nella sintesi di acido 9-cis retinoico.
4.12 Determinazione dei livelli di 9-cis retinolo nella frazione microsomale e citosolica e suo possibile ruolo quale precursore del 9-cis RA
Avendo dimostrato la capacità esclusiva della via biosintetica “solubile” di impiegare quale substrato 9-cis retinolo per la sintesi di acido 9-cis retinoico, abbiamo ritenuto importante, al fine di evidenziare un possibile ruolo fisiologico di tale attività enzimatica, verificare la presenza, o una possibile via di formazione del 9-cis retinolo in frazioni microsomali e citosoliche isolate da fegato di ratto o bue.Per valutare la possibilità che il 9-cis retinolo si formi nelle cellule attraverso il processo di mobilizzazione dei retinil esteri nei microsomi, sfruttando l’energia liberata durante l’idrolisi, si è studiato l'effetto della CRBPI e di altre proteine leganti i retinoidi sul processo di idrolisi dei retinil esteri, la forma principale di accumulo della Vitamina A all'interno dei tessuti. Questo esperimento è stato effettuato anche per valutare l'importanza del trasportatore proteico nel veicolare il retinolo verso specifici siti di metabolizzazione intracellulare, agendo da modulatore di particolari attività enzimatiche. L’attività enzimatica RE idrolasica (REH), presente nella frazione microsomale, non è stata ancora isolata, ma è già stato dimostrato l'effetto stimolatorio della CRBP in forma apo nella reazione di idrolisi dei retinil esteri e conseguente formazione di oloCRBP (Ottonello et al. 1987; Boerman e Napoli, 1991).Esperimenti effettuati incubando microsomi ottenuti da fegato di bue o ratto in presenza o in assenza di un eccesso di apoCRBPI (10 M), hanno evidenziato un significativo incremento della attività retinil estere idrolasica quando la proteina legante il retinolo era aggiunta alla miscela di reazione (Tabella 17). L'incremento della attività RE-idrolasica basale indotto dalla proteina legante in forma apo (circa 2 volte) è risultato essere altamente specifico per la CRBPI.
Aggiunta SpecieRetinolo (pmoli)
Nessuna Ratto 83
ApoCRBPI Ratto 185
ApoCRBPII Ratto 93
Nessuna Bue 50
ApoCRBPI Bue 103
ApoCRBPII Bue 52
63

Risultati
Tabella 17: Idrolisi dei retinil esteri. Le reazioni sono state incubate a 37°C per 30’, pH 8, 150 mM KCl, in presenza di 700 µg di proteine microsomali, e, quando presente 10 M di CRBPI o II.
La CRBPII, un’altra proteina legante il retinolo localizzata esclusivamente nell'intestino, che presenta una identità del 55% con la CRBP e pressochè totale sovrapponibilità strutturale, non incrementa significativamente l'attività idrolasica, suggerendo così l'esistenza di un selettivo riconoscimento della CRBPI da parte del sistema enzimatico.E' stato inoltre analizzato lo stato, libero o legato, e la forma isomerica del retinolo mobilizzato nella reazione di idrolisi stimolata da apoCRBPI (Tabella 18). In modo analogo a quanto osservato precedentemente in presenza di apoCRBPI, la quantità totale di retinolo recuperato è circa 2 volte maggiore rispetto a quanto ottenuto quando non è aggiunta la apoCRBPI. In quest’ultimo caso il retinolo mobilizzato dalla reazione basale viene quasi interamente recuperato nella frazione sedimentata contenente i microsomi (94%-), mentre è presente prevalentemente nel surnatante solubile (87%) quando le miscele di reazione contengono un eccesso di apoCRBPI. La frazione solubile ottenuta dopo incubazione con la apoCRBPI presenta uno spettro di assorbimento UV-visibile identico a quello del complesso all-trans retinolo-CRBPI. Ciò è confermato dalla analisi in HPLC della forma isomerica del retinolo idrolizzato, che è esclusivamente nella configurazione all-trans.
Frazione Apo CRBPIRetinolo (pmoli)
Olo CRBPI
Microsomi - 56 -
Solubile - 3 -
Microsomi + 16 -
Solubile + 116 +
Tabella 18: Localizzazione e forma isomerica del retinolo idrolizzato. Le reazioni sono state incubate nelle condizioni indicate in Tabella 17, in presenza di 500 µg di proteine microsomali di ratto, e, quando presente 10 M di CRBPI.
Quest'ultimo dato è piuttosto importante, e contrariamente a quanto accade per la formazione di 11-cis retinolo nell'epitelio pigmentato retinico, dove l'energia derivante dall'idrolisi dei retinil esteri può essere impiegata per favorire l'isomerizzazione di all- trans a 11-cis retinolo (Nagao e Olson, 1994), esclude la possibilità che l'isomero 9-cis del retinolo possa generarsi in seguito ad idrolisi dei retinil esteri, rendendosi così disponibile per la sintesi di acido 9-cis retinoico.Avendo eliminato la possibilità che i microsomi, ricchi di retinil esteri, costituiscano una fonte importante per la sintesi di 9-cis retinolo, è stata verificata la presenza di tale
64

Risultati
isomero quale componente endogeno dei citosol da fegato di ratto e bue utilizzati per la sintesi di RA. Una indicazione positiva in questo senso costituirebbe infatti una importante evidenza in favore del ruolo fisiologico della via “solubile” di sintesi di 9-cis RA da precursori aventi configurazione isomerica 9-cis. Le rispettive frazioni solubili sono state quindi analizzate per la presenza di differenti forme isomeriche di retinolo.L’estrazione in solvente organico e la successiva analisi in HPLC ha evidenziato la presenza di retinolo in citosol freschi sia di ratto che di bue (Tabella 19), ma non in frazioni da citosol bovino precedentemente frazionate su gradienti di glicerolo (140 e 220 kDa). La configurazione isomerica del retinolo recuperato nei vari citosol è risultata essere esclusivamente all-trans. Inoltre, incubando i citosol in condizioni di reazione (2 mM NAD, 37°C, pH 8) per 30’, è stata rilevata la sintesi di acido retinoico, esclusivamente nella forma all-trans. La formazione di RA dipende in modo lineare dalla quantità di proteina impiegata, è drasticamente inibita dall’aggiunta di apoCRBPI 5 M, e non è rilevabile, nelle medesime condizioni, con il componente da citosol di bue di 140 kDa.
FrazioneRetinolo endogeno
(pmoli)Acido Retinoico
sintetizzato (pmoli)
Citosol Ratto 10 7
Citosol Bue 6 3.5
Citosol Ratto + apoCRBPI 10 N.D.
140 kDa Bue N.D. N.D.
Tabella 19: Rilevazione della quantità di retinolo endogeno e reattività in varie frazioni subcellulari. Sono stati analizzati 400 g di proteina, e quando presente, apoCRBPI alla concentrazione di 5 M. Le reazioni sono state incubate in condizioni standard per 30’ a 37°C. I valori sono tutti stati normalizzati per l’efficienza di estrazione
Questi dati, sebbene evidenzino la presenza e la reattività “RA biosintetica” di retinolo nella forma all-trans quale componente endogeno dei citosol di fegato di ratto e bue, non forniscono elementi a favore dell’ipotesi che il 9-cis retinolo possa rappresentare precursore in vivo per la sintesi di acido 9-cis retinoico. Non escludono tuttavia che il 9-cis retinolo possa avere un importante ruolo fisiologico nella sintesi di acido 9-cis retinoico: è possibile, ad esempio, che le condizioni sperimentali impiegate per lo studio dell’idrolisi dei retinil esteri fossero inadeguate per la formazione di tale composto, oppure che i livelli di 9-cis retinolo presenti nei vari citosol esaminati fossero al di sotto del limite di rivelazione.
65

Risultati
4.13 Parziale purificazione e caratterizzazione dell’attività solubile da fegato di ratto
Al fine di isolare e meglio caratterizzare il componente enzimatico “solubile” presente nel citosol di fegato di ratto, ottenere maggiori informazioni riguardo alla presenza di distinti componenti molecolari del sistema, e lo stato dell’intermedio di reazione retinaldeide, un estratto epatico solubile (125 mg di proteina citosolica) è stato frazionato mediante filtrazione molecolare su una colonna Sephacril S-300 HR, e le diverse frazioni sono state saggiate per l’attività retinoico biosintetica.Rispetto all’analisi precedentemente effettuata su citosol bovino mediante sedimentazione su gradiente di glicerolo, ci si è avvalsi in questo caso della possibilità di impiegare quale substrato differenti forme isomeriche di retinolo e retinaldeide, oltre ad una tecnica di analisi in HPLC ad alta risoluzione ed estremamente rapida, che ci ha permesso di identificare e quantificare contemporaneamente differenti isomeri di retinaldeide e acido retinoico.Per i saggi di sintesi di acido retinoico si sono impiegati come substrati all-trans e 9-cis retinolo sia in forma libera (10 M) che legata alla CRBPI (4 M), in presenza di 2 mM NAD come cofattore.Come illustrato in Figura 26 A, con tutti i substrati impiegati il picco di attività acido retinoico biosintetica è stato individuato in corrispondenza della frazione 64, avente un peso molecolare apparente di 140 kDa, in buon accordo con quanto già identificato in citosol bovino. La concomitante determinazione dell’intermedio di reazione nelle frazioni testate per la sintesi di acido retinoico ci ha permesso di rilevare l’accumulo di quantità cospicue di retinaldeide, con un massimo nella frazione 68 (avente un peso molecolare apparente di circa 80 kDa), ben distinto da quello determinato nel caso dell’attività Ra biosintetica.La dipendenza della sintesi di RA dalla presenza di cofattori nelle frazioni isolate dopo gel filtrazione mostra la medesima selettività per il NAD già individuata nel citosol non frazionato: l’attività RA biosintetica in presenza di NADP, è risultata essere dal 80 all 85% inferiore a quella misurata impiegando NAD, sia utilizzando 9-cis che all-trans retinolo, in forma libera o legata a CRBP quale substrato. Le frazioni isolate da citosol di ratto sono state quindi saggiate per la loro capacità di ossidare la retinaldeide ad acido retinoico, fornendo quale substrato all-trans e 9-cis retinaldeide: in entrambi i casi il massimo di attività RA biosintetica è stato identificato in corrispondenza della frazione 58, avente un peso molecolare approssimativo di 220 kDa, che presumibilmente identifica l’attività retinaldeide deidrogenasica del sistema “misto” (Figura 26 B). L’attività specifica relativa all’ossidazione del retinolo e della retinaldeide, rispettivamente nelle frazioni 64 e 58 è risultata essere da 5 a 12 volte maggiore di quella determinata nel citosol non frazionato.
66

Risultati
Title: D:\CACCA\FIG12.PSCreator: GSS*CGI PostScript DriverCreationDate: 4 February 1996 18:18
Figura 26: Frazionamento del citosol di fegato di ratto mediante cromatografia di gel filtrazione. (A) Sintesi di RA e RALD da all-trans e 9-cis retinolo (10 M) incubati in presenza di un volume costante (25 ml) delle frazioni eluite e 2 mM NAD come cofattore. Il medesimo profilo è stato ottenuto incubando gli stessi retinoidi (4 M) in forma legata a CRBPI. (B) Sintesi di RA da all-trans e 9-cis retinaldeide (10 M) incubati nelle medesime condizioni indicate in (A).
Al contrario, impiegando all-trans e 9-cis retinolo come substrato sotto forma di complesso con la CRBPI, non si è osservato nessun incremento significativo dell’attività specifica RA biosintetica, e ciò è presumibilmente dovuto all’effetto inibitorio associato
67

Risultati
alla apoCRBPI generata durante il processo ossidativo. Abbiamo cercato di ottenere maggiori informazioni riguardo al possibile rilascio nel mezzo di reazione o al trasferimento diretto all’interno del sistema enzimatico dell’intermedio 9-cis retinaldeide, effettuando un esperimento di competizione, tra all-trans retinaldeide e 9-cis retinolo in presenza della frazione 64.I dati ottenuti (Tabella 20) indicano l’esistenza di una effettiva competizione tra 9-cis e all-trans retinaldeide, e suggeriscono che l’intermedio di ossidazione diffonda liberamente nel mezzo di reazione. L’assenza di un meccanismo di trasferimento diretto dell'intermedio retinaldeide all'interno del sistema enzimatico di ossidazione sarebbe in buon accordo con quanto riportato da Kim et al. (Kim et al., 1992), che hanno eseguito un analogo esperimento di competizione in citosol di fegato di ratto tra all-trans retinolo marcato radioattivamente e all-trans retinaldeide.
Prodotto
Substrato 9-cis RA
(pmoli)
All-trans RA
(pmoli)
9-cis RALD
(pmoli)
All-trans RALD
(pmoli)
9-cis ROH 62 0 50 0
All-trans RALD 0 23 72 394
9-cis ROH +
All-trans RALD
4.5 21 94 379
Tabella 20: Competizione tra 9-cis ROH e all-trans RALD. L’esperimento è stato eseguito incubando in condizioni standard di reazione la frazione 64 (25 g di proteina totale) in presenza di 9-cis ROH (10 M), e/o all-trans RALD (5.3 M) (cofattore NAD 2 mM).
L’analisi della conversione di vari retinoidi in frazioni da citosol di fegato di ratto isolate mediante gel filtrazione, ha quindi rilevato la presenza di un massimo di attività RA biosintetica nella frazione 64 (140 kDa), frapposta ad altri due picchi di attività, retinaldeide e retinolo deidrogenasica presenti nelle frazioni 58 (220 kDa) e 68 (80 kDa). L’attività acido retinoico biosintetica presente nella frazione 64 potrebbe essere costituita da un unico componente enzimatico, oppure dall’intersezione di due componenti distinti presenti nelle frazioni 58 e 68.Per distinguere tra queste due possibilità, abbiamo valutato l’effetto sulla sintesi di RA del mescolamento di frazioni contenenti la massima attività retinolo (# 58) e retinaldeide deidrogenasica (# 68), rispetto all’attività presente nella sola frazione 64. I dati riportati in Tabella 21 mostrano la presenza di una effettiva complementazione tra le attività retinolo e retinaldeide deidrogenasica presenti nelle frazioni 58 e 68 per la sintesi di RA, che, pur essendo maggiore di quella presente nella sola frazione 64, non aumenta in modo sinergico in seguito alla presenza simultanea delle due frazioni.
68

Risultati
n° FrazioneProteina totale
(g)Acido retinoico
(pmoli)Retinaldeide
(pmoli)
58 16 0 0
64 25 67 38
68 16 3 64
58+68 16 77 30
58+68 32 101 47
Tabella 21: Sintesi di RA e RALD in differenti frazioni eluite da gel filtrazione. Le reazioni sono state condotte in presenza di 9-cis retinolo (10 M) quale substrato, più 2 mM NAD, per 30’ a 37°C, utilizzando un volume costante (20 l) di frazioni, con l’eccezione dell’ultimo saggio in cui il volume totale di frazioni utilizzato è di 40 l.
Sebbene il risultato dell’esperimento sia interessante, non permette di discriminare con chiarezza quale delle due ipotesi presentate sia valida.Dal momento che l’eventuale dimostrazione dell’esistenza di due distinte retinolo deidrogenasi (una in grado di formare direttamente RA nella frazione 64, l’altra di sintetizzare RALD nella 68) avrebbe escluso l’ipotesi che la sintesi di RA nella frazione 64 fosse causata dall’intersezione delle due attività ossidasiche presenti nelle frazioni 58 e 68, abbiamo analizzato secondo differenti criteri la capacità retinolo ossidasica delle frazioni 64 e 68 al fine di identificare un possibile comportamento differenziale.A questo proposito, è stato inizialmente analizzato l’effetto del pirazolo, uno specifico inibitore delle alcool deidrogenasi, sulla formazione di RA e RALD da parte delle due retinolo deidrogenasi.
n° FrazioneAttività residua (%)
Acido retinoico Retinaldeide
64 18 14
68 28 16
140 kDa 68 N.D.
Tabella 22: Inibizione dell’attività deidrogenasica di frazioni da citosol di bue e ratto in presenza di 1 mM pirazolo. Le reazioni sono state incubate in condizioni standard di reazione, impiegando 10 M 9-cis retinolo quale substrato, e un volume costante (20 l) della frazione proteica.
69

Risultati
In presenza di concentrazioni di pirazolo (1mM) che inibiscono totalmente l’attività etanolo deidrogenasica delle ADH presenti nel citosol di fegato di ratto non frazionato, e influiscono in modo modesto sull’attività retinoico biosintetica della frazione 140 kDa isolata da fegato bovino, abbiamo osservato una significativa inibizione, quantitativamente simile nelle frazioni 64 e 68 da citosol di ratto, della sintesi di RA e RALD (Tabella 22).Il medesimo comportamento è stato osservato impiegando retinolo in forma legata a CRBPI quale substrato. La cospicua inibizione dell’attività osservata in entrambe le frazioni testate, fornisce un elemento a favore dell’esistenza di un’unica attività retinolo deidrogenasica, la quale potrebbe appartenere alla famiglia delle alcool deidrogenasi.Anche l’effetto inibitorio sulla sintesi “solubile” di RA indotto dalla presenza di microsomi è stato studiato per evidenziare differenze nel comportamento delle frazioni 64 e 68. Analogamente a quanto osservato con la frazione citosolica cruda, l’aggiunta di minime quantità di microsomi da fegato di ratto alla miscela di reazione inibisce drasticamente (oltre 80%) la sintesi sia di RA che di RALD, sia nella frazione 64 che nella 68, a partire da 9-cis retinolo (10 M) quale substrato.Nessuno dei dati ottenuti dall’esperimento di mescolamento delle frazioni, la sensibilità al pirazolo, ed alla aggiunta di microsomi, ha permesso di acquisire elementi che permettono di differenziare tra le attività retinolo deidrogenasiche presenti nelle frazioni 64 e 68. L’ipotesi più probabile rimane pertanto quella che spiega l’esistenza dell’attività acido retinoico biosintetica nella frazione 64 con la presenza di due attività complementanti, una retinolo e una retinaldeide deidrogenasi, entrambe NAD-dipendenti, derivate dalle “code” dei picchi di attività delle frazioni 58 e 68. La possibilità che l’attività RA biosintetica “solubile” presente in citosol di fegato di ratto, potenzialmente importante per la sintesi in vivo dell’acido 9-cis retinoico, sia costituita da un unico componente enzimatico, oppure da due distinte attività (un’alcool deidrogenasi in grado di ossidare il retinolo, e la medesima retinaldeide deidrogenasi presente nella via “mista”), potrà essere chiarita in seguito all’ulteriore isolamento e purificazione dell’attività enzimatica solubile mediante altri metodi cromatografici.
4.14 Biosintesi di acido retinoico in cellule somatiche di testicolo di ratto
E’ noto che la vitamina A e l’acido retinoico svolgono un ruolo essenziale nel processo di spermatogenesi e nel mantenimento dell’epitelio germinale dei tubuli seminiferi: in ratti nutriti con una dieta priva di vitamina A la maturazione delle cellule germinali si arresta all’inizio del processo di meiosi (Eskild e Hansson, 1994). La spermatogenesi è ristabilita dalla somministrazione di retinolo, o da iniezioni intraperitoneali di acido retinoico. Una dieta ricca di RA non è in grado di restaurare le normali condizioni in quanto tale molecola non è in grado di attraversare la barriera emato-testicolare, selettivamente permeabile, che è formata dalle cellule somatiche del testicolo, le Sertoli e le peritubulari.
70

Risultati
Esse circondano l’epitelio seminifero nutrendolo e proteggendolo, e cooperano nell’assorbimento di micronutrienti dal sangue, così come alla formazione della matrice intercellulare.In entrambi i tipi cellulari delle cellule somatiche del testicolo sono stati identificati vari tipi di RAR (Kim e Griswold, 1990; Eskild et al., 1991; Huang et al., 1994), e sono presenti livelli relativamente elevati di CRBPI (Porter et al., 1985), mentre la CRABPI è presente solo nelle cellule del Sertoli (Blaner et al., 1987). L’espressione della CRBPI e di altri prodotti importanti per il mantenimento della corretta funzione testicolare, è indotta nelle cellule del Sertoli dall’attivazione trascrizionale dei recettori nucleari da parte di RA.Dal momento che il contenuto di RA nel testicolo è pressochè indipendente dai livelli plasmatici di questo retinoide, la sua sintesi in situ riveste presumibilmente una notevole importanza per il mantenimento dei processi dipendenti dalla vitamina A in quest’organo, che costituisce così un interessante modello per lo studio della sintesi in vitro di RA. Inoltre, a differenza del fegato, le cellule somatiche del tubulo seminifero sono tutte disponibili sotto forma di colture primarie, arricchite nei diversi tipi cellulari.Poichè non è stato ancora chiarito se, e in quale dei diversi tipi cellulari del testicolo, avvenga la sintesi di RA, presumibilmente a partire dal complesso retinolo-CRBP, abbiamo testato, con la collaborazione della professoressa Michela Galdieri dell’Università di Roma, che ha isolato i differenti tipi cellulari, la capacità RA biosintetica delle cellule somatiche del testicolo di ratto, con la speranza che questo tipo di analisi ci fornisse inoltre altri elementi per valutare il significato dell’esistenza di due distinti sistemi acido retinoico biosintetici.Il saggio in vitro di biosintesi di RA è stato applicato inizialmente su frazioni isolate da testicoli interi di ratti, al fine verificare preliminarmente la presenza e la reattività dei due sistemi di biosintesi di acido retinoico, e confrontare i valori di attività con quelli osservati in fegato. Sono state scelte condizioni di reazione analoghe a quelle impiegate per le precedenti analisi, e una concentrazione di oloCRBPI fisiologica, ma ampiamente superiore a quella delle Km identificate nei sistemi biosintetici di fegato (Tabella 23).
La sintesi di RA è stata rilevata in entrambi i sistemi enzimatici impiegando frazioni subcellulari da testicolo di 20 giorni di età, e l’attività retinoico biosintetica della via “mista” è risultata essere sensibilmente maggiore (di circa 12 volte) rispetto a quella del sistema “solubile”, a differenza di quanto avviene nel fegato di animali della stessa età, dove l’attività rilevata dei due sistemi è analoga. L’attività retinoico biosintetica del sistema “misto” in testicolo totale è circa il 30% maggiore di quella osservata in fegato di animali della stessa età, a differenza del sistema “solubile”, attivo circa il 15% rispetto a quello di fegato. Esperimenti di sintesi di RA a partire da all-trans retinolo in forma libera (5 M) hanno evidenziato nel sistema “solubile” di fegato di ratto e bue l’esistenza del medesimo rapporto di attività (il fegato risulta circa 6 volte più attivo) osservato in precedenza impiegando oloCRBPI come substrato.
71

Risultati
OrganoSistema
enzimaticoAcido Retinoico
(pmol/ 30’)
Fegato Solubile 22
Fegato Misto 26
Testicolo Solubile 3
Testicolo Misto 38
Tabella 23: Sintesi di RA in frazioni subcellulari di testicolo e fegato di animali di 20 giorni. Le reazioni sono state eseguite incubando 200 µg di proteine citosoliche e/o 200 µg di proteine microsomali a 37°C per 30’ in presenza di 2 mM NAD e/o NADP e 4 M olo-CRBPI quale substrato.
Avendo accertato la presenza dell’attività retinoico biosintetica dei due sistemi enzimatici, abbiamo cercato di localizzare in quale dei tipi cellulari del testicolo fosse presente, testando la reattività di frazioni subcellulari delle cellule somatiche.La sintesi in vitro di acido retinoico è stata individualmente saggiata in frazioni subcellulari da cellule peritubulari e del Sertoli di ratti di 20 giorni di vita; come mostrato in Tabella 24, le cellule peritubulari non sono attive nemmeno impiegando le condizioni di reazione più favorevoli (quando è presente esclusivamente oloCRBPI quale substrato), mentre nelle cellule del Sertoli l’attività retinoico biosintetica è rilevabile in tutte le condizioni di reazione nel caso del sistema “misto”, ed è presente nel sistema “solubile”
Tipo cellulare Apo CRBPIFrazione
subcellulareAcido Retinoico
(pmoli/30’)
Sertoli - Citosol 2.3
Sertoli - Citosol-Microsomi 16.2
Sertoli + Citosol N.D.
Sertoli + Citosol-Microsomi 5.8
Peritubulari - Citosol N.D.
Peritubulari - Citosol-Microsomi N.D.
Tabella 24: Sintesi di RA in frazioni subcellulari di differenti tipi cellulari di testicolo da animali di 20 giorni. Le reazioni sono state eseguite incubando 200 µg di proteine citosoliche e/o 200 µg di proteine microsomali a 37°C per 30’ in presenza di 2 mM NAD e/o NADP e 4 M olo-CRBPI.
72

Risultati
Title: D:\CACCA\FIG13.PSCreator: GSS*CGI PostScript DriverCreationDate: 4 February 1996 18:23
Figura 27: Analisi mediante complementazione eterologa della capacità acido retinoico biosintetica di frazioni subcellulari di cellule peritubulari. Le reazioni sono state incubate impiegando 4 M olo e apo CRBPI, e 2 mM ciascuno NAD e NADP (A) Microsomi di cellule del Sertoli (150 g), e citosol di frazioni eterologhe (200 g) sono impiegati come fonte di attività biosintetica. I valori riportati sono stati ottenuti dopo sottrazione della quota di RA sintetizzata dai soli microsomi in presenza di cofattori e substrato (1.5 pmoli) (B) Citosol di cellule del Sertoli (200 g) è stato incubato con microsomi eterologhi (150 g) e testato per la sintesi di RA. I valori determinati sono sottratti per la quota di RA sintetizzata dai soli microsomi, pari a 1.5, 0 e 2.5 pmoli per i microsomi di Sertoli, di peritubulari e di fegato di ratto, rispettivamente.
73

Risultati
solo in assenza di apoCRBPI, ma risulta otto volte meno attiva. Come rivelato da esperimenti di controllo, l’attività delle frazioni cellulari isolate direttamente da testicolo non è diversa da quelle coltivate per 5 giorni in vitro.L’assenza di qualsiasi attività acido retinoico biosintetica in cellule peritubulari è sorprendente, considerato che queste cellule sono attivamente coinvolte nell’internalizzazione del retinolo, e contengono concentrazioni relativamente elevate di oloCRBPI, ma è in linea con quanto osservato recentemente da Davis e Ong (Davis e Ong, 1995), che non rilevano la presenza di alcuna attività in grado di esterificare il retinolo della oloCRBPI in microsomi di cellule peritubulari.Abbiamo cercato di confermare questo importante dato, cercando di identificare con un esperimento di complementazione eterologa quale delle frazioni di cellule peritubulari, citosolica, microsomale, o entrambe, fosse responsabile della assenza di attività di sintesi di acido retinoico osservata nel sistema “misto”. L’attività retinoico biosintetica di frazioni citosoliche o microsomali di cellule peritubulari di 20 giorni è stata testata in combinazione con la frazione complementare da cellule del Sertoli.Come mostrato in Figura 27, in condizioni in cui una effettiva complementazione per l’attività RA biosintetica era osservata tra frazioni di fegato di ratto e di cellule del Sertoli, praticamente nessuna sintesi di RA era rilevata da citosol o microsomi di cellule peritubulari in presenza della frazione complementare da cellule del Sertoli. Questo dato implica che l’assenza di un’attività rilevabile del sistema “misto” in cellule peritubulari è imputabile all’assenza di entrambe le attività retinolo e retinaldeide deidrogenasica.Poichè l’attività retinoico biosintetica identificata nel sistema “solubile” di cellule del Sertoli di animali di 20 giorni è risultata essere al di sotto dei limiti di rivelazione in presenza di concentrazioni equimolari di apo e olo CRBPI (Tabella 24), che rappresentano condizioni “fisiologiche” per le cellule del Sertoli (Kato et al., 1985), abbiamo proceduto con una serie di esperimenti mirati ad ottimizzare e a caratterizzare l’attività dell’unica via biosintetica potenzialmente in grado anche in vivo di sintetizzare RA: il sistema “misto“.In presenza di concentrazioni equimolari di olo e apoCRBPI, e di un quantità costante di proteina citosolica, la sintesi di RA nel sistema “misto” delle cellule del Sertoli aumenta all’aggiunta di proteina microsomale, fino alla saturazione in presenza di un rapporto 1:2 micromsomi:citosol (Figura 28 A).L’intermedio retinaldeide è sempre rilevabile, e si accumula in presenza di un eccesso di microsomi, suggerendo così che il secondo passaggio ossidativo sia quello limitante nella sintesi di RA.La sintesi di RA è massima a pH 8, e risponde in modo lineare all’incremento della proteina citosolica e microsomale così come alla dipendenza del tempo di incubazione (fino a 90 minuti) (dati non mostrati).
74

Risultati
Title: D:\CACCA\FIG14.PSCreator: GSS*CGI PostScript DriverCreationDate: 4 February 1996 18:24
Figura 28: Caratterizzazione della sintesi di RA nel sistema “misto” di cellule del Sertoli. (A) Sintesi di RA e RALD a partire da concentrazioni equimolari di olo e apoCRBPI (4 M), in presenza di una quantità costante (200 g) di proteine citosoliche, e quantità crescenti di microsomi. (B) Dipendenza dai cofattori della sintesi di RA e RALD. NAD e NADP sono stati impiegati ad una concentrazione di 2 mM. (C) Sintesi di RA in presenza di concentrazioni crescenti di una miscela equimolare di olo e apoCRBPI. Incubazione a 37°C per 45’. (D) Analisi in HPLC della configurazione isomerica dell’acido retinoico sintetizzato da cellule del Sertoli nel sistema “misto”. Una identica distribuzione isomerica è stata osservata in seguito incubazione di all-trans RA in condizioni di reazione senza cofattori e successiva estrazione. Tutte le reazioni sono effettuate in condizioni standard di reazione, impiegando 200 g di proteina citosolica e 100 g di proteina microsomale di frazioni isolate da animali di 20 giorni.
La produzione di RA in cellule del Sertoli di animali di 20 giorni dipende dalla presenza dei cofattori NAD e NADP, in modo analogo a quanto già identificato precedentemente per il sistema ”misto“ in fegato e bue. Infatti, come mostrato nella Figura 28 B, in presenza di NAD e NADP la sintesi di RA è massima, ma quantità rilevabili di RA e RALD sono presenti anche in presenza dei singoli cofattori, in modo tale da confermare l’impiego preferenziale da parte della RoDH microsomale di NADP, e di NAD dalla RalDH citosolica. I dati riportati in Figura 28 C mostrano che la sintesi di RA nel sistema “misto” di cellule del Sertoli è saturabile, ed è massima in presenza di circa 4 M di oloCRBPI, concentrazione analoga a quella stimata (1-9 M) in cellule del Sertoli (Cavazzini et al., 1996), e la Vmax apparente (circa 73 pmol/h mg di proteina citosolica) è
di circa tre volte minore di quella ottenuta in assenza di apoCRBPI.
75

Risultati
La forma isomerica dell’acido retinoico sintetizzato in cellule del Sertoli è stata determinata (Figura 28 D): analogamente a quanto osservato nel sistema “misto“ di fegato di bue o di ratto impiegando quale substrato all-trans retinolo-CRBPI, l’acido retinoico prodotto in vitro in queste condizioni è recuperato esclusivamente nella configurazione all-trans.Il risultato di questi esperimenti di caratterizzazione e ottimizzazione dei vari parametri di reazione del saggio di sintesi di RA in cellule del Sertoli da animali di 20 giorni, che non hanno rilevato evidenti differenze rispetto ai parametri già noti per il sistema “misto” da fegato di ratto e bue, consente di affermare che i due componenti enzimatici della via di sintesi “mista“, precedentemente identificati in fegato di ratto e bue, sono presenti nelle frazioni microsomale e citosolica di cellule del Sertoli.Poichè l’espressione di CRBPI e CRABPI, così come quella dei recettori nucleari per l’acido retinoico, mostrano caratteristici e distinti andamenti temporali di espressione (Eskild e Hansson, 1994; Rajan et al., 1990; Schmitt e Ong, 1993), ci siamo chiesti se, e in quale modo, l’attività acido retinoico biosintetica dipendesse dall’età degli animali. Come riportato in Figura 29, la sintesi aumenta di circa 5 volte tra il 12° e il 20° giorno di vita, e si mantiene poi pressochè costante sia in presenza che in assenza di apoCRBPI. Nelle stesse condizioni, la produzione di RA nel sistema “misto” in fegato di ratto mostra
Title: D:\CACCA\FIG15.PSCreator: GSS*CGI PostScript DriverCreationDate: 4 February 1996 18:28
Figura 29: Livelli di attività acido retinoico biosintetica in cellule del Sertoli isolate da animali di differenti età. La sintesi di RA nel sistema “misto” di cellule del Sertoli è stata testata incubando 4 M oloCRBPI presenza (barre a strisce) o in assenza (barre bianche) di concentrazioni equimolari di apoCRBPI. I dati riportati come 34/SCOC sono stati ottenuti in condizioni standard di reazione (200 + 100 g di citosol e microsomi), da cellule del Sertoli ottenute da animali di 34 giorni, nelle quali le cellule germinali erano state rimosse mediante trattamento iposmotico selettivo prima del frazionamento.
76

Risultati
un andamento sostanzialmente differente, rimanendo costante fino a 37 giorni, per aumentare di circa due volte nell’animale adulto (dato non mostrato).L’incremento osservato nella sintesi di RA nelle cellule del Sertoli tra il 12° e il 20° giorno potrebbe essere spiegato con un incremento del fabbisogno di acido retinoico in concomitanza dell'inizio della meiosi, processo che comincia attorno al ventesimo giorno di vita dell'animale. Considerato l’incremento della popolazione di cellule germinali, che si accrescono in concomitanza con l’inizio della meiosi, potrebbe essere possibile spiegare l'incremento di attività misurata in cellule del Sertoli con la presenza di cellule germinali contaminanti le colture.Tale ipotesi è stata tuttavia esclusa dall’osservazione che, in coincidenza con il continuo incremento della popolazione di cellule germinali che avviene tra il 20° e il 37° giorno di vita dell’animale, l’attività RA biosintetica rimane essenzialmente costante, e dal fatto che non è stata osservata nessuna differenza significativa di attività retinoico biosintetica tra colture cellulari "normali" isolate da animali di 34 giorni e altre colture deprivate delle cellule germinali mediante trattamento osmotico selettivo (Figura 29).Questi dati localizzano quindi per la prima volta un’attività di sintesi di acido retinoico in uno specifico tipo cellulare del testicolo di ratto. I dati a nostra disposizione hanno escluso la presenza di un’attività retinoico biosintetica in cellule peritubulari, e, indirettamente, nelle cellule germinali del tubulo seminifero. Le cellule del Sertoli, importanti nel supportare la crescita e il differenziamento degli spermatozoi, si sono dimostrate le uniche in grado di ossidare il complesso retinolo-CRBP attraverso il sistema biosintetico misto in condizioni compatibili con la concentrazione intracellulare di CRBPI, ed è pertanto assai probabile che esse svolgano un ruolo importante anche nella biosintesi in vivo di acido retinoico.
77

Discussione
5. DISCUSSIONE
La messa a punto di un metodo di dosaggio in vitro dell’attività acido retinoico biosintetica rapido e versatile, che può utilizzare retinoidi dispersi in solvente organico o il complesso retinolo-CRBPI in concentrazioni fisiologiche come substrati, e frazioni subcellulari citosoliche o microsomali provenienti da diversi tessuti animali come fonte di attività enzimatica, ha costituito la premessa necessaria per lo studio e l’analisi di questo importante processo della farmacocinetica della vitamina A. Rispetto al precedente saggio di attività che impiegava [3H] retinolo-CRBP bovina, ora oloCRBPI ricombinante di ratto è stata utilizzata come substrato; la nuova procedura di analisi prevede inoltre l’impiego di diversi possibili metodi di estrazione in solvente organico dei retinoidi, e di differenti e specifiche fasi mobili da noi individuate per l’analisi dei prodotti mediante HPLC, che consentono di dosare velocemente e con elevata sensibilità differenti isomeri di retinolo, retinaldeide ed acido retinoico.L’analisi dell’attività acido retinoico biosintetica in differenti organi animali impiegando il nuovo saggio “a freddo” ha rilevato la presenza nelle frazioni subcellulari isolate da fegato di bue e ratto di entrambi i sistemi di biosintesi di RA, la via “solubile” e quella “mista”. La versatilità e la elevata sensibilità del saggio ci ha inoltre permesso di testare per la prima volta la capacità RA biosintetica di differenti tipi cellulari del testicolo di ratto, organo nel quale la formazione di acido retinoico è indispensabile per il mantenimento della spermatogenesi.I dati raccolti hanno confermato preliminarmente la presenza nell’organo intero di un’attività enzimatica in grado di sintetizzare acido retinoico. Il tentativo di identificare il sito di produzione di questo composto mediante l’analisi della capacità biosintetica dei principali tipi cellulari presenti nel tubulo seminifero del testicolo di ratto, ha evidenziato la presenza di tale attività esclusivamente nelle frazioni subcellulari isolate da cellule del Sertoli. In condizioni di reazione fisiologiche (in presenza di concentrazioni M di olo e apoCRBPI) il solo sistema biosintetico attivo per la produzione di acido retinoico nelle cellule del Sertoli è risultato essere quello “misto”, e il prodotto è stato recuperato esclusivamente nella configurazione all-trans.E’ noto da tempo il ruolo essenziale nel testicolo delle cellule del Sertoli, le quali, secernendo numerosi composti biologicamente attivi, mantengono nel tubulo seminifero il microambiente ottimale che consente la corretta proliferazione e il differenziamento delle cellule germinali a spermatozoi. I dati ottenuti suggeriscono l’esistenza di un nuovo importante ruolo per queste cellule, che potrebbero quindi essere, anche in vivo quelle fisiologicamente deputate alla biosintesi di acido retinoico nel testicolo.Una attenta caratterizzazione biochimica delle attività biosintetiche “solubile” e “mista” presenti nelle frazioni isolate da organi di bue e ratto, ci ha permesso di identificare alcune importanti differenze tra le due, che ci hanno consentito di formulare alcune
78

Discussione
ipotesi riguardo il possibile ruolo fisiologico e il significato dell’esistenza di due sistemi enzimatici che risultano apparentemente in grado di catalizzare la medesima reazione.La via biosintetica “mista” di ratto e bue è in grado di impiegare quale substrato all-trans retinolo in forma libera o sotto forma di complesso con la CRBPI per sintetizzare esclusivamente acido all-trans retinoico. La selettività di questo sistema, a differenza di quello “solubile”, riguardo la capacità di impiegare come substrato differenti forme isomeriche di retinoidi è notevole: il 9-cis retinolo non viene riconosciuto come substrato dalla retinolo deidrogenasi presente in microsomi di fegato di ratto e bue, il che è in accordo con quanto recentemente osservato da Boerman e Napoli (Boerman e Napoli, 1995) impiegando l’enzima in forma parzialmente purificata, a differenza di quanto avviene con la retinaldeide deidrogenasi solubile, che è invece in grado di ossidare ad acido retinoico all-trans e 9-cis retinaldeide. Questo dato esclude un qualsiasi ruolo del sistema “misto” nella sintesi dell’acido 9-cis retinoico a partire da precursori aventi la stessa configurazione isomerica, quali ad esempio il 9-cis retinolo.I parametri cinetici ottenuti impiegando quale substrato all-trans retinolo-CRBPI nel sistema “misto” (la Km è perfettamente compatibile con la concentrazione endogena di tale proteina), i dati che evidenziano l’impiego preferenziale di NADP e NAD rispettivamente da parte della RoDH microsomale e dalla RalDH solubile, così come il peso molecolare stimato di quest’ultimo componente del sistema “misto” (220-230 kDa) ricavato in seguito a parziale purificazione in citosol di bue e ratto, sono in buon accordo con quanto precedentemente riportato in letteratura (Napoli, 1994).Il confronto tra la reattività del sistema “misto” di ratto con quello di bue ha evidenziato una maggiore produzione di RA da parte di quest’ultimo di circa 10 volte nelle stesse condizioni sperimentali. Un riuscito esperimento di complementazione eterologa ha dimostrato che la minore velocità di sintesi di RA osservata nel sistema “misto” di ratto è in realtà causata dalla scarsa reattività del componente solubile, la retinaldeide deidrogenasi NAD-dipendente, mentre risulta invece sostanzialmente identica l’attività retinolo deidrogenasica NADP-dipendente presente nei microsomi delle due specie. La minore reattività della RalDH del sistema “misto” di ratto costituisce quindi presumibilmente la causa della presenza di una maggiore quantità di retinaldeide rilevata come intermedio di reazione quando viene testato il sistema “misto” di ratto rispetto a quando vengono impiegate frazioni subcellulari isolate da fegato di bue.In presenza di un eccesso di apoCRBPI, impiegata per simulare al meglio la condizioni intracelluari, la sintesi di RA nel sistema “misto” a partire da all-trans retinolo-CRBPI , seppure inibita, è ben rilevabile, a differenza di quanto accade nel sistema “solubile”. Questo dato, così come i precedenti, per quanto ottenuti mediante esperimenti effettuati in vitro, supporta l’ipotesi che il sistema “misto” possa essere quello fisiologicamente deputato, anche in vivo, alla sintesi di acido all-trans retinoico impiegando oloCRBPI quale substrato.
79

Discussione
Tra l’altro, alcuni dei dati ottenuti evidenziano come l’espressione dei componenti enzimatici che costituiscono il sistema “misto”, possa essere modulata nei vari tessuti o tipi cellulari in funzione dell’età dell’animale per rispondere ad un incrementato (o diminuito) fabbisogno di RA: ad esempio, mentre non sono rilevabili differenze nell’attività retinoico biosintetica del sistema “misto” in fegato di ratti fino a 34 giorni di vita, l’aumentata richiesta di RA nel testicolo di animali di circa 20 giorni di età, coincidente con l’inizio della meiosi delle cellule germinali, potrebbe essere la causa del sensibile incremento di attività acido retinoico biosintetica rilevato nel sistema “misto” di cellule del Sertoli tra il 12° e il 20° giorno di vita dell’animale.La maggior parte dei risultati riportati nel presente lavoro di tesi, nonostante siano state ottenute nuove ed importanti informazioni riguardo le due attività che costituiscono il sistema “misto” di bue e ratto, riguarda la caratterizzazione biochimico-funzionale e la parziale purificazione del sistema “solubile” di sintesi di acido retinoico, identificato per la prima volta nel nostro laboratorio nella frazione solubile di fegato bovino, che, a differenza di quello “misto”, non era stato ancora isolato ed identificato nei suoi componenti molecolari.I risultati ottenuti hanno evidenziato la presenza anche nel citosol di fegato di ratto di un’attività deidrogenasica capace di ossidare in vitro all-trans retinolo, disperso in solvente organico o sotto forma di complesso stechiometrico con la CRBPI, ad acido all-trans retinoico.Il confronto tra la reattività del sistema “solubile” di bue e ratto per la produzione di acido retinoico, a differenza di quanto osservato per quello “misto”, non presenta differenze significative. Ma l’accumulo dell’intermedio di reazione retinaldeide nel sistema “solubile” è minimo o nullo quando sono impiegati rispettivamente all-trans retinolo in forma libera o oloCRBPI in bue, mentre risulta chiaramente rilevabile in presenza di entrambi i substrati nel citosol di fegato di ratto.Le attività “solubili” di fegato di ratto e bue presentano altre differenze: una riguarda la selettività nell’impiego del coenzima, poiché, mentre NAD è l’unico cofattore impiegato dal citosol di fegato di ratto, anche NADP, seppure con minore efficienza, può essere utilizzato dalla frazione solubile di bue per sintetizzare RA. Inoltre, anche il curioso e tuttora non spiegato effetto inibitorio dei microsomi sulla sintesi di RA nel sistema “solubile” è stato rilevato esclusivamente in ratto.Queste differenze avanzano alcuni dubbi circa l’identità delle due attività “solubili” identificate in ratto e bue, ma l’analisi per gel filtrazione o mediante gradiente di glicerolo ha mostrato come queste possiedano il medesimo peso molecolare apparente (140 kDa).Quando è impiegata come substrato CRBPI in forma olo legante esclusivamente all-trans retinolo in concentrazioni fisiologiche, l’attività RA biosintetica presente nel citosol di fegato di ratto e bue è drasticamente inibita dalla presenza di un eccesso di apoCRBPI ( @ 5% di attività residua in presenza di concentrazioni equimolari di apo e olo CRBPI).
80

Discussione
Nonostante la curva di inibizione osservata si avvicini a quella attesa nell’ipotesi che la causa sia il sequestramento del retinolo, non è comunque possibile escludere sulla base dei dati a nostra disposizione, che l’effetto osservato sia in realtà causato da un’azione diretta della apoCRBPI sull’enzima ossidante, analogamente a quanto avviene nel caso di LRAT, dove la oloCRBPI è substrato, e la apoCRBPI è un potente inibitore competitivo (Herr e Ong, 1992).Differenti curve di inibizione da apoCRBPI sono state osservate nel sistema “solubile” di bue impiegando come substrato oloCRBPI bovina o ricombinante di ratto contenenti piccole percentuali di 9-cis retinolo, rispetto a quelle riportate in questo lavoro di tesi, ottenute con nuovi preparati di all-trans retinolo-CRBPI (cfr. Figure 11 e 30). Per cercare di chiarire i motivi di questo differente comportamento, l’esperimento è stato ripetuto impiegando come substrato la oloCRBPI ricombinante di ratto impiegata precedentemente, ma analizzando questa volta i prodotti con un metodo di eluizione in HPLC che permette di identificare la forma isomerica dell’acido retinoico sintetizzato.
Title: D:\CACCA\FIG11.PSCreator: GSS*CGI PostScript DriverCreationDate: 4 February 1996 15:55
Figura 30: Inibizione da apoCRBPI sulla sintesi di RA nel sistema “solubile” in citosol di fegato bovino. In condizioni standard di reazione, e in presenza di 400 g di citosol, oloCRBPI (4 M) è incubata in presenza di concentrazioni crescenti di apoCRBPI ricombinante di ratto.
Come mostrato in Figura 30, la curva di inibizione ottenuta in presenza di concentrazioni crescenti di apoCRBPI, che riporta la formazione di acido retinoico in citosol di fegato bovino senza differenziare le differenti specie isomeriche presenti ( l ), è simile a quella precedentemente ottenuta (vedi Figura 11). Ma completamente differenti tra loro sono le curve di inibizione relative alle singole specie isomeriche di RA recuperate nelle medesime condizioni di reazione: la sintesi di all-trans RA ( ) viene drasticamente
81

Discussione
inibita già in presenza di concentrazioni equimolari di apoCRBPI, mentre quella di 9-cis RA ( o ) lo è solo in modo molto modesto, tanto che risulta la sola forma isomerica prodotta in presenza di un eccesso di apoCRBPI. L’andamento osservato è spiegabile se si assume che siano presenti in realtà entrambe le forme isomeriche all-trans e 9-cis del retinolo in forma legata alla oloCRBPI, le quali sono entrambe reattive nel sistema solubile, ma diversamente sensibili alla presenza della apoCRBPI, in modo analogo a quanto osservato quando sono state ottenute le curve di inibizione impiegando come substrato differenti forme di CRBP leganti esclusivamente all-trans o 9-cis retinolo (Cfr Figure 30, 23 e 24). In realtà, quando è stato effettuato per la prima volta questo esperimento, le tecniche a disposizione non permettevano la separazione delle differenti forme isomeriche di acido retinoico prodotto: il 9-cis RA era appena stato scoperto essere l’attivatore trascrizionale di RXR, i dati presenti in letteratura riportavano l’assoluta specificità di legame della CRBPI per l’all-trans retinolo, ed era perciò pressoché impossibile interpretare in modo differente da quanto fatto i dati ottenuti.Sebbene non sia stato possibile chiarire con certezza il meccanismo inibitorio della apoCRBPI, ma assumendo che questo non sia un artificio presente esclusivamente nei saggi in vitro, questi dati suggeriscono quindi che la sintesi in vivo di all-trans RA da parte del sistema “solubile” possa avvenire quindi solamente quando siano presenti concentrazioni intracellulari minime o nulle di apoCRBPI.Sebbene sia difficile escludere con certezza un possibile ruolo fisiologico del sistema “solubile” per la formazione di RA sulla base di esperimenti eseguiti in vitro, il confronto della reattività dei due sistemi biosintetici in condizioni di reazione che simulano quelle “fisiologiche” (in presenza di all-trans retinolo-CRBPI e apoCRBPI), suggerisce che la sintesi di acido all-trans retinoico anche in vivo, nei vari tessuti, sia catalizzata dal sistema “misto”, l’unico attivo in presenza di un eccesso di apoCRBPI. Ma altri risultati da noi ottenuti suggeriscono un altro importante ruolo per il sistema “solubile” di fegato di ratto e bue: potrebbe essere responsabile della formazione dell’acido 9-cis retinoico, molecola essenziale per numerosi processi biologici, la cui origine non è ancora stata chiarita.Abbiamo verificato sperimentalmente le principali ipotesi proposte riguardo la formazione dell’acido 9-cis retinoico: inizialmente è stata analizzata la possibile formazione di quest’ultimo in seguito ad isomerizzazione di acido all-trans retinoico, catalizzata da proteine citosoliche o microsomali, o dalla presenza di apoCRABPI. Ma la minima percentuale di 9-cis RA recuperato (3% max.) in seguito ad incubazione di all-trans RA, indipendente dalla quantità di proteina impiegata, o dalla presenza o meno di apoCRABPI, tende ad escludere che tale processo possa avere una qualche importanza fisiologica. Risulta infatti difficile ipotizzare che la sintesi di una molecola ad elevata attività biologica come il 9-cis RA non sia catalizzata da un’attività enzimatica. Tra l’altro, 9-cis RA modula l’attività trascrizionale di tutti i principali recettori nucleari, tra i quali i RAR, leganti l’acido all-trans retinoico. Sembra piuttosto improbabile che all-
82

Discussione
trans RA, molecola con un tempo di emivita estremamente rapido, la cui sintesi enzimatica e i livelli intracellulari sono strettamente regolati, possa anche essere il precursore di 9-cis RA, che ha numerosi ed importanti “bersagli” intracellulari, e necessita quindi presumibilmente di un controllo rigoroso ed indipendente della sintesi e del mantenimento dei livelli fisiologici.Altri esperimenti ci hanno consentito di verificare, analizzando la configurazione isomerica dell’acido retinoico formato nelle vie biosintetiche “solubile” e “mista” a partire dal complesso fisiologico all-trans retinolo-CRBPI, l’eventuale presenza di una deidrogenasi-isomerasi in grado di sintetizzare 9-cis RA. Anche in questo caso, il risultato sperimentale ha escluso la possibilità che un enzima con queste caratteristiche fosse presente, poichè sia il sistema “misto” che quello “solubile” presenti in fegato e testicolo di o bue mantengono la configurazione isomerica all-trans del retinolo, sintetizzando, con velocità diverse, esclusivamente all-trans RA. Nel corso di questi esperimenti, la verifica della forma isomerica del retinolo legato ai vari pool di CRBPI impiegati, ha evidenziato la presenza costante di una piccola percentuale (3%) di 9-cis retinolo, che ci ha suggerito la possibile esistenza di un’interazione specifica tra questo retinoide e la CRBPI. In effetti, nonostante dati precedenti riportati in letteratura, riportassero l’assenza di binding, è stato possibile dimostrare con una differente tecnica di analisi che la CRBPI lega con elevata affinità (Kdiss: 10-8 M) tale composto, ed isolare così per la prima volta il complesso 9-cis
retinolo-CRBPI.La reattività di quest’ultimo, o del 9-cis retinolo in forma libera è stata testata in entrambi i sistemi, “misto” e “solubile”, di ratto e bue, al fine di verificare la validità dell’ipotesi della sintesi di 9-cis RA da precursori aventi la stessa configurazione isomerica. Mentre il sistema “misto” non è in grado di ossidare il 9-cis retinolo, il sistema “solubile” di ratto e bue converte efficientemente tale composto, sia in forma libera che legata a CRBPI, formando come prodotto 9-cis RA, anche in presenza di concentrazioni fisiologiche di apoCRBPI.Diversi esperimenti sono stati eseguiti per cercare di chiarire se, nel sistema “solubile” di ratto e bue, quando sono incubati differenti retinoidi sotto forma di complesso con la CRBPI, l’enzima ossidante riconosca come substrato il retinolo presente in equilibrio, oppure, come evidenza di specificità, esista un riconoscimento specifico tra enzima ossidante e proteina trasportatrice in forma olo.Se si considera che i valori di Km ottenuti impiegando come substrato retinoidi in forma
libera dispersi in solvente organico sono di circa 10-5 M (7 M per l’ossidazione di all-trans retinolo nel caso del sistema ”solubile” di bue), la possibilità di convertire efficacemente in vitro concentrazioni di circa 10-8 / 10-7 M di retinolo libero in equilibrio con la oloCRBPI (5 M) risulta piuttosto remota.Non si può comunque escludere un effetto drastico sui valori dei parametri cinetici relativi all’ossidazione dei retinoidi in forma libera causato dalla presenza del solvente
83

Discussione
(DMSO o etanolo) necessario per solubilizzare i retinoidi: il reale valore di Km per la
conversione dei retinoidi in equilibrio con la CRBPI in assenza del solvente organico potrebbe essere in realtà notevolmente inferiore rispetto a quello determinato quando è presente.A questo proposito è interessante ricordare il dato ottenuto riguardo la reattività di all-trans retinolo endogeno nei citosol di ratto e bue. Assumendo che non siano stati commessi grossolani errori nella stima della quantità di retinolo, e considerato che non conosciamo in quale forma esso sia presente (è comunque sensibile alla dialisi), la sintesi di all-trans RA, che aumenta linearmente in base alla quantità di proteina impiegata, avverrebbe in presenza di concentrazioni di retinolo dell’ordine di 10-7 M, circa 100 volte inferiore ai valori di Km ottenuti per la conversione di all-trans retinolo libero disperso in
DMSO nel citosol di bue.E’quindi estremamente difficile e delicato identificare condizioni sperimentali che permettano di distinguere con chiarezza se avvenga il riconoscimento del complesso o del retinolo libero da parte dell’attività enzimatica: è comunque possibile, impiegando rapporti costanti tra apo e olo CRBPI, creare condizioni di reazione che, teoricamente, mantengono pressochè costante la concentrazione di retinolo libero in equilibrio, aumentando la concentrazione della olo proteina. In questo caso, impiegando il complesso 9-cis retinolo-CRBPI quale substrato, si è osservata sintesi di 9-cis RA con un andamento chiaramente saturabile, indice di un riconoscimento preferenziale della oloCRBPI da parte dell’attività ossidante, piuttosto che del retinolo presente in equilibrio.Ma è anche possibile variare esclusivamente la concentrazione di retinolo in equilibrio mantenendo costante la concentrazione di oloCRBPI: occorre incubare concentrazioni crescenti di apoCRBPI in presenza di una concentrazione costante di olo proteina. In questo caso, impiegando 9-cis retinolo-CRBPI come substrato, si osserva un effetto inibitorio sulla sintesi di 9-cis RA in presenza di concentrazioni crescenti di apoCRBPI, ma, analogamente a quanto osservato nel caso del sistema “misto” in presenza di all-trans retinolo-CRBPI, la curva di inibizione osservata non correla apparentemente con quella teorica di inibizione attesa nel caso fosse riconosciuto il retinolo presente in equilibrio. Occorre comunque ricordare ancora come curve di inibizione teoriche sensibilmente differenti possano essere costruite assumendo valori leggermente differenti di Kdiss.
Futuri esperimenti potranno contribuire a stabilire con certezza quale sia il reale meccanismo con il quale la apoCRBPI esercita il suo effetto inibitorio.Considerando i dati nel loro complesso, si può quindi affermare che la sintesi di 9-cis RA impiegando concentrazioni crescenti di 9-cis retinolo-CRBPI è saturabile, sembra dipendere dalla concentrazione del complesso, suggerendo l’esistenza di un possibile specifico riconoscimento da parte della retinolo deidrogenasi del sistema “solubile”, e presenta parametri cinetici che sono analoghi a quelli in precedenza ottenuti in reazioni nelle quali era coinvolto il complesso all-trans retinolo-CRBPI. Ovviamente non sono ancora disponibili dati riguardo l’esistenza in vivo e la concentrazione di tale complesso
84

Discussione
che permettano di valutare il possibile significato fisiologico di tale reazione. Diverse evidenze in letteratura riportano comunque la presenza di 9-cis retinolo in diversi tessuti animali, sebbene nessun trasportatore specifico, plasmatico o intracellulare sia stato ancora identificato. Abbiamo quindi verificato se 9-cis retinolo potesse formarsi come prodotto del processo di idrolisi dei retinil esteri stimolato da apoCRBPI, oppure fosse presente, come componente endogeno nei citosol delle frazioni solubili da noi impiegate. In entrambi i casi, esclusivamente retinolo nella forma all-trans è stato individuato, quale prodotto della reazione retinil estere idrolasica , in forma legata alla CRBPI, oppure come componente endogeno presente nel citosol di fegato di ratto e bue. Nel caso della reazione di idrolisi dei retinil esteri, è stato comunque confermato l’effetto stimolatorio della apoCRBPI in ratto, ed evidenziato per la prima volta la presenza di tale reazione anche nei microsomi di bue, con formazione di oloCRBPI come prodotto. Anche se non è stato ancora spiegato il meccanismo stimolatorio della apoCRBPI, è importante notare che, in presenza della CRBPII, proteina avente una struttura terziaria pressochè sovrapponibile alla proteina di tipo I, ma una minore affinità per l’all-trans retinolo, non si è rilevato alcun incremento dell’attività idrolasica, in modo analogo a quanto avviene con l’enzima microsomale LRAT, che è inibito dalla CRBPI, ma non dalla CRBPII.quando è stata invece verificata la presenza nella frazione solubile di 9-cis retinolo, che i dati in letteratura riportano essere presente in vivo in concentrazioni pari a circa il 10% di quelle della forma all-trans, è possibile che tale composto non sia stato rilevato perchè presente nelle frazioni analizzate in quantità inferiori al limite di rilevazione dell’analisi in HPLC. La mancanza di riferimenti certi riguardo la presenza e la concentrazione fisiologica di 9-cis retinolo in forma libera o legata a CRBPI nei tessuti, rende incerta la valutazione del possibile ruolo anche in vivo del sistema “solubile” di ratto e bue nella sintesi di 9-cis RA; sebbene alcuni dati siano stati riportati precedentemente in letteratura, risulta questa la prima chiara evidenza sperimentale che identifica una possibile via metabolica responsabile dell’origine del 9-cis RA. Nuove ed importanti informazioni riguardo i componenti molecolari che costituiscono il sistema “solubile”, confrontabili con quelle precedentemente raccolte impiegando fegato di bue nel tentativo di isolare l’attività solubile su di un gradiente di glicerolo, sono state ottenute con il frazionamento del citosol di fegato di ratto mediante gel filtrazione per dimensioni molecolari. L’impiego del saggio di attività RA biosintetico nelle frazioni eluite in seguito a gel filtrazione ha evidenziato la presenza di un picco di attività avente un peso molecolare apparente di circa 140000 (analogo a quello in precedenza identificato in bue), in grado di ossidare all-trans e 9-cis retinolo in forma libera o legati a CRBPI in modo strettamente dipendente dalla presenza di NAD come cofattore, analogamente a quanto osservato nel citosol non frazionato. L’impiego di all-trans e 9-cis retinaldeide come substrato ha identificato un picco di attività acido retinoico biosintetica ben distinto dal
85

Discussione
precedente, non in grado di ossidare retinolo, avente un peso molecolare apparente di circa 220000, che è presumibilmente, per dimensioni molecolari e specificità di substrato, l’attività retinaldeide deidrogenasica solubile del sistema “misto”. Anche in questo caso, il peso molecolare di questo componente è simile a quello precedentemente identificato in citosol da fegato bovino.L’impiego di una nuova procedura di analisi in HPLC dei prodotti, che permette di rilevare contemporaneamente RA e l’intermedio di reazione, non disponibile quando era stato frazionato il citosol di bue, ha evidenziato l’accumulo di retinaldeide nella frazione avente peso molecolare apparente di 80000 impiegando retinolo come substrato. I successivi esperimenti hanno cercato di chiarire se l’attività del sistema “solubile”, la RoDH di 140 kDa fosse in realtà costituita da un singolo o due componenti aventi le stesse dimensioni, oppure dall’intersezione delle “code” delle attività RoDH di 80 kDa e della RalDH del sistema “misto”. Quest’ultima ipotesi è resa possibile dall’osservazione che le attività delle frazioni di 80 e 220 kDa, le quali singolarmente non sono in grado di produrre acido retinoico a partire da retinolo, sono in grado di complementare tra loro per formare RA quando vengono riunite.Un esperimento di competizione tra retinolo e retinaldeide, effettuato impiegando la frazione di 140 kDa, ha dimostrato che l’intermedio viene rilasciato nel mezzo di reazione, rendendo così meno probabile la presenza di un’unica attività capace di catalizzare direttamente la sintesi di RA da ROH, e suggerendo invece come più probabile l’esistenza di due distinte attività che catalizzano ognuna uno step della reazione di sintesi di acido retinoico, analogamente a quanto avviene nel sistema “misto”. Questa osservazione è in accordo con quanto riportato da Kim et al. (Kim et al., 1992), che rilevano l’esistenza di una effettiva competizione tra retinaldeide e [3H] retinolo in citosol di ratto non frazionato.Nel tentativo di escludere l’ipotesi dell’intersezione delle “code” dei picchi per spiegare la presenza di una attività RA biosintetica di 140 kDa, diversi esperimenti hanno tentato, senza peraltro riuscirvi, di rilevare differenze e di dimostrare che sono funzionalmente distinte sia le attività RalDH presenti nella frazione di 140 kDa e di 220 kDa, così come le retinolo deidrogenasi individuate individuate nelle frazioni di 140 e 80 kDa.In particolare, si è tentato di meglio caratterizzare l’attività retinolo deidrogenasica seguendo la formazione di acido retinoico e retinaldeide da retinolo nelle frazioni di 140 kDa e 80 kDa, che risulta in entrambe pressochè totalmente inibita dall’aggiunta di minime quantità di proteine microsomali, o dalla presenza di 1 mM pirazolo, uno specifico inibitore delle ADH, in modo analogo a quanto osservato da Kim et al. nel citosol di ratto non frazionato (Kim et al., 1992).L’insieme dei dati raccolti, pur senza fornire alcun elemento che escluda in modo definitivo la possibile presenza di un’unica attività RA di 140 kDa nelle frazioni isolate da fegato di ratto, è in ottimo accordo con l’ipotesi che prevede la presenza
86

Discussione
contemporanea nella frazione di peso molecolare apparente 140000 delle “code” delle
All-trans RALD
CITOPLASMA
9-cis RA
oloCRBPI + apoCRBPI
9-cis retinolo
MICROSOMIRoDH
(34 kDa) NADP+
9-cis RALD
NAD+ ADH (?)(80 kDa)
All-trans RA
RalDH (220 kDa)
NAD+
NAD+
All-trans retinolo
Figura 31: Schema delle possibile vie di sintesi di 9-cis e all-trans RA in citosol e microsomi di fegato di ratto.
deidrogenasi presenti nelle frazioni di 80 e 220 kDa, il che suggerisce una importante conclusione. Il sistema “solubile” di fegato di ratto sarebbe quindi costituito in realtà da due distinti componenti, una retinolo e una retinaldeide deidrogenasi NAD-dipendenti (Figura 31). Come suggerito dalle dimensioni molecolari e dalla notevole sensibilità al pirazolo, il primo componente del sistema “solubile” potrebbe essere un enzima appartenente alla classe I delle alcool deidrogenasi in grado di ossidare 9-cis e all-trans
87

Discussione
retinolo sia in forma libera che legata alla CRBPI, mentre il secondo componente, la retinaldeide deidrogenasi, sulla base del peso molecolare apparente e della capacità di ossidare sia 9-cis che all-trans retinaldeide, potrebbe essere la stessa attività che catalizza il secondo step di ossidazione nel sistema “misto”. Ulteriori esperimenti di caratterizzazione delle frazioni isolate, ma soprattutto un’ulteriore frazionamento e purificazione delle frazioni isolate, potranno confermare, o confutare, queste conclusioni.Anche il sistema “solubile” da fegato bovino è costituito da due attività enzimatiche distinte? Il confronto tra i dati raccolti frazionando il citosol di fegato di bue su gradiente di glicerolo, sebbene incompleti a causa del differente tipo di analisi eseguita, con quelli ottenuti in ratto, presenta alcune analogie e differenze. I pesi molecolari apparenti della RoDH (130 kDa) e RalDH (230 kDa) bovine sono simili a quelli a quelli delle corrispondenti attività identificate in ratto, ma differente è la sensibilità mostrata al pirazolo dalla frazione di 130 kDa, la cui attività RA biosintetica è scarsamente inibita. Inoltre, analogamente a quanto accade nel citosol non frazionato, quando è impiegata la frazione di 130 kDa da bue in presenza di oloCRBPI come substrato, a differenza di quella di ratto di 140 kDa, non è rilevabile la presenza dell’intermedio retinaldeide, anche se non sono disponibili dati riguardo il suo possibile accumulo nelle altre frazioni.E’possibile interpretare la differente sensibilità della frazione bovina di 130 kDa al pirazolo e l’assenza di retinaldeide come un’indicazione che i sistemi “solubili” di ratto e bue sono in realtà completamente distinti, ma occorre tenere presente che la classe IV di alcool deidrogenasi è presente in fegato, ed è scarsamente sensibile al pirazolo.Inoltre, è già stata dimostrata la maggiore reattività della RalDH del sistema “misto” di bue, fino a 10 volte più attiva di quella presente in citosol di ratto. Se anche il sistema “solubile” di bue fosse costituito da due componenti distinti (una ADH e la RalDH del sistema “misto”), sarebbe facilmente spiegabile la causa del minore accumulo di retinaldeide osservato rispetto a quanto avviene nel sistema “solubile” di ratto, con la maggiore reattività mostrata dalla retinaldeide deidrogenasi del sistema “misto” bovino.Sulla base di questa ipotesi, è quindi possibile che i vari tentativi di purificazione del componente solubile di bue non abbiano avuto buon esito perchè, avendo rilevato l’assenza di retinaldeide, il mancato sinergismo nella sintesi di RA quando erano riuniti i picchi laterali, e la mancata competizione della retinaldeide nei confronti di [3H] retinolo-CRBPI nel citosol non frazionato, si era ipotizzato che fosse presente un unico componente nella frazione 140 kDa da citosol di fegato bovino. In realtà, quest’ultimo poteva essere costituito da due distinti componenti, la cui separazione, e non l’inattivazione e l’instabilità dell’attività isolata, poteva essere la causa reale della scarsa o nulla attività RA biosintetica recuperata in presenza di retinolo come substrato durante i differenti tentativi di ulteriore purificazione dell’attività “solubile”.
88

Conclusioni
6. CONCLUSIONI
I risultati riportati nel presente lavoro di tesi hanno fornito nuovi elementi per la conoscenza dei meccanismi che sono alla base dell’attivazione metabolica intracellulare della vitamina A. Il momento più importante della farmacocinetica del retinolo è senz’altro costituito dalla sua ossidazione ad acido retinoico, il retinoide biologicamente più attivo.E’ stato quindi sviluppato e realizzato un saggio in vitro che permette di dosare in modo rapido, sensibile e versatile l’attività acido retinoico biosintetica delle due vie di sintesi “mista” e “solubile” finora identificate nelle frazioni subcellulari di vari tessuti animali, impiegando il substrato naturale all-trans retinolo-CRBPI, e altri retinoidi in forma libera o legata al trasportatore intracellulare del retinolo. E’ stato inoltre possibile dimostrare per la prima volta l’esistenza di uno specifico legame tra il 9-cis retinolo, un possibile precursore per la sintesi di 9-cis RA, e la CRBPI, ed il complesso così formato è stato isolato ed impiegato come substrato nei saggi di attività.Questo metodo di dosaggio dell’attività retinoico biosintetica da noi messo a punto, potrà anche essere sfruttato per la ricerca di variazioni ditale attività in cellule provenienti da tessuti normali, patologici, o trattati con retinoidi. Una prima interessante applicazione in questo senso è stato il tentativo di acquisire informazioni riguardo alla presenza e la localizzazione dell’attività retinoico biosintetica nel testicolo di ratto, organo nel quale l’acido retinoico svolge un ruolo essenziale per il mantenimento della spermatogenesi.Tra i vari tipi cellulari del tubulo seminifero esaminati, le cellule del Sertoli sono risultate le sole in grado di catalizzare, in condizioni di reazione fisiologiche, la sintesi di acido all-trans retinoico impiegando il substrato naturale all-trans retinolo-CRBPI, esclusivamente attraverso la via biosintetica “mista”. La caratterizzazione biochimica delle due vie di sintesi “solubile” e “mista” di acido retinoico presenti in frazioni subcellulari di fegato di ratto e di bue ha suggerito l’esistenza di distinti possibili ruoli in vivo per queste due attività enzimatiche. L’analisi della configurazione isomerica dell’acido retinoico prodotto ha evidenziato la capacità del sistema "misto" di ratto o bue, di sintetizzare acido all-trans retinoico in condizioni fisiologiche, impiegando un substrato naturale quale all-trans retinolo-CRBPI; i dati hanno mostrato inoltre la notevole stereoselettività di tale sistema nei confronti dell'isomero all-trans, legato alla CRBP o in forma libera, e l'incapacità di utilizzare 9-cis retinolo quale substrato. Quest’ultimo retinoide, in forma libera o come complesso con la CRBP, viene invece efficientemente ossidato ad acido 9-cis retinoico dal sistema “solubile”, che è anche in grado di catalizzare, in assenza dell’inibitore apoCRBPI, la sintesi di all-trans RA da all-trans retinolo in forma libera o legata alla CRBPI. Sebbene rimanga da accertare la presenza ed il ruolo in vivo di retinoidi precursori aventi configurazione 9-cis e del complesso 9-cis retinolo-CRBPI, l’attività solubile individuata
89

Conclusioni
in citosol di ratto e bue rappresenta il primo caso di sintesi enzimatica in vitro dell’acido 9-cis retinoico.Il frazionamento per dimensioni molecolari del citosol di fegato di ratto e bue ha permesso per la prima volta di ottenere importanti informazioni sui componenti molecolari che costituiscono il sistema “solubile”. I risultati ottenuti suggeriscono l’esistenza di due attività enzimatiche, entrambe NAD-dipendenti, delle quali la prima, di circa 80 kDa, potrebbe essere un enzima appartenente alla famiglia delle alcool deidrogenasi, in grado di catalizzare la sintesi di 9-cis e all-trans retinaldeide dalle corrispondenti forme isomeriche di retinolo, mentre la seconda è una retinaldeide deidrogenasi solubile di circa 230 kDa, che potrebbe essere lo stesso enzima che nel sistema “misto” catalizza l’ossidazione della retinaldeide generata dalla retinolo deidrogenasi NADP-dipendente presente nella frazione microsomale. I risultati qui presentati forniscono quindi nuovi elementi che contribuiscono a chiarire i complessi processi metabolici che portano alla formazione delle differenti forme isomeriche di acido retinoico in grado di regolare l’espressione di importanti geni mediante il legame con i recettori nucleari RAR e RXR, e permettono di acquisire le conoscenze di base sulle caratteristiche funzionali e sui componenti molecolari dei sistemi acido retinoico biosintetici che potranno essere sfruttate in futuro per la regolazione in vivo a scopo terapeutico della biosintesi intracellulare di acido retinoico.
90

Materiali e Metodi
7. MATERIALI E METODI
7.1 Conservazione e stabilità dei retinoidi
I retinoidi in polvere sono stabili per circa un anno se conservati in essiccatore, in atmosfera controllata di azoto, al buio e a 4 °C. La medesima stabilità si è osservata in seguito a conservazione a -80 °C. Una volta solubilizzati in metanolo o in etanolo, i retinoidi sono conservati al buio a -20° C, e non mostrano significativa isomerizzazione per un periodo di almeno 2 mesi, ad eccezione dei vari isomeri della retinaldeide che restano stabili solo per alcuni giorni. La concentrazione dei retinoidi va verificata ogni volta prima del loro utilizzo poichè in alcuni casi può diminuire significativamente (10% 25% circa dopo 30 giorni nel caso del retinolo e della retinaldeide, rispettivamente). Occorre mantenere sempre in ghiaccio e al buio le soluzioni durante il loro utilizzo, ed è sempre consigliabile, vista la notevole instabilità di questi composti, preparare al momento le soluzioni da utilizzare.I coefficienti di estinzione molare (cm-1.M-1) utilizzati per la determinazione della concentrazione dei vari retinoidi, e la lunghezza d’onda alla quale deve essere effettuata la misura spettrofotometrica in soluzione etanolica, sono indicati in Tabella 25.
CompostoCoefficiente di estinzione molare
(cm-1.M-1)
Lunghezza d’onda
(nm)
Acido all-trans retinoico 45000 350
Acido 9-cis retinoico 36500 343
Acido 13-cis retinoico 39800 354
All-trans retinolo 46000 325
9-cis retinolo 36930 325
13-cis retinolo 42170 325
11-cis retinolo 30450 325
all-trans retinaldeide 43200 383
9-cis retinaldeide 35000 383
Tabella 25: Coefficienti di estinzione molare di vari retinoidi
7.2 Espressione e purificazione di CRBPI e CRBPII ricombinante da Escherichia coli in forma apo e olo
L'espressione e la purificazione di CRBPI e II di ratto è stata ottimizzata apportando alcune modifiche alla procedura descritta da Levin et al. (Levin et al., 1990); il metodo
91

Materiali e Metodi
qui riportato, pressochè identico per la CRBPI e II, permette la purificazione ad omogeneità di entrambe le proteine grazie a due rapidi passaggi cromatografici.Il plasmide pMON contenente il gene codificante per la CRBPI o II di ratto posto sotto il controllo di un promotore inducibile da acido nalidixico, ed un sito che conferisce la resistenza all’ampicillina, è stato utilizzato per trasformare un ceppo di batteri E.coli JM101.Crescita delle colture batteriche ed espressione della CRBPDopo crescita overnight, 20 ml di cellule batteriche vengono inoculate in 1 l di terreno LB (Luria Broth) contenente 0.1 mg/ml di ampicillina, ed incubate a 30 °C.Quando i batteri raggiungono un valore di OD600 di 0.2 (dopo circa 1 ora e 15 minuti)
viene aggiunto l’induttore (0.05 mg/ml di acido nalidixico, stock 10 mg/ml in 0.1N NaOH) che, attivando il promotore recA del plasmide, permette la trascrizione del gene della CRBP di ratto.Le cellule batteriche sono incubate per circa 3 ore e 30 minuti a 30 °C fino a raggiungere un valore di OD600 di 0.85-1.0, quindi vengono centrifugate a 9000g per 10 minuti al
fine di raccogliere le cellule, che vengono poi risospese in 2-3 volumi di tampone di lisi (50 mM Tris, pH 7.9; 10% saccarosio, 0.5 mM PMSF)Lisi dei batteriTutte le operazioni seguenti, di lisi batterica e i vari passaggi di purificazione della proteina sono eseguite a 4 °C. La lisi viene effettuata mediante sonicazione: otto colpi di 15 secondi con pause di circa 1 minuto. La formazione di schiuma e la variazione di densità della soluzione indicano l’avvenuta rottura delle cellule batteriche.Purificazione della CRBPI e IISi centrifuga l’omogenato a 9000g per 10' e si preleva il surnatante. A questo si aggiunge lentamente un uguale volume di una soluzione satura di solfato d'ammonio contenente DTT (15 mM), e si lascia in agitazione blanda per circa 2 ore. Si centrifuga quindi a 41000g per 30’ e si preleva il liquido surnatante nel quale si trova la CRBP.Nel caso si purifichi la CRBPII, al surnatante ottenuto viene aggiunto nuovamente solfato d’ammonio per raggiungere il 70% di saturazione, si lascia quindi in agitazione per 2 ore, e si centrifuga poi con le stesse modalità descritte in precedenza.Il precipitato così ottenuto nel caso della CRBP II, o il surnatante contenente il 50% del solfato d’ammonio nel caso si purifichi la CRBPI vengono dializzati overnight contro 5 litri di tampone di colonna (25 mM potassio fosfato pH 7.2, 200 mM NaCl , 3 mM ß-mercaptoetanolo). In alternativa si può abbassare la conducibilità dei campioni fino a circa 800µS mediante diluizione successiva in cella Amicon con tampone di colonna. Nel caso si purifichi CRBP in forma olo, si satura la proteina con un eccesso 1.5 molare di all-trans retinolo solubilizzato in etanolo, preparato al momento, per minimizzare la formazione di differenti forme isomeriche.La purificazione di proteine saturate con retinoidi mediante colonne cromatografiche rende queste ultime non utilizzabili successivamente per la purificazione di apo-proteine.
92

Materiali e Metodi
Infatti, i retinoidi trattenuti dalla matrice, per quanto in minima quantità, vengono legati con elevata affinità dalle proteine, che, caricate in forma apo, eluiscono in forma parzialmente saturata (si è osservato fino al 15% di saturazione con la olo-CRBP). Occorre quindi utilizzare colonne distinte per la purificazione di proteine in forma apo e in forma olo.Il campione, (max.volume: 4% volume di colonna) viene caricato su di una colonna a filtrazione molecolare ULTROGEL Aca54, ad un flusso di 0.5 ml/min. L’eluizione della CRBP (raccolta in frazioni di 7.5 ml), viene individuata, se la proteina è in forma olo, misurando la fluorescenza (l di eccitazione = 350 nm, l di emissione = 485 nm), oppure mediante analisi dello spettro d’assorbimento UV-visibile, che presenta un picco caratteristico a 350 nm. Nel caso si purifichi proteina in forma apo, questa può essere identificata saturando un’aliquota delle frazioni eluite con retinolo e analizzando poi lo spettro di assorbimento.Dopo questo passaggio cromatografico, la purezza della proteina (stimata dall’analisi dello spettro di assorbimento della olo-CRBP, o mediante elettroforesi) è stata, in differenti preparazioni, di 70-95% nel caso della CRBPI , e di oltre il 90% per la CRBPII. Nel caso si intenda purificare ad omogeneità CRBPI contenente 100% retinolo nella forma all trans, è consigliabile saturare la proteina solo prima del successivo passaggio cromatografico.Per purificare ad omogeneità la proteina, si applica il campione ad una colonna a scambio anionico QMA (gruppo funzionale metilamina quaternaria) mediante FPLC.La soluzione proteica contenente CRBP viene equilibrata con il tampone di colonna (25 mM Tris, pH 7.3) per ridurre la salinità (10-15 µS max.).Una volta caricato il campione sulla colonna ad un flusso di 1.5 ml/min, viene applicato un gradiente lineare (flusso 2.5 ml/min) da 0 a 200 mM NaCl in 40’, e vengono raccolte frazioni di 1.5 ml.L’eluizione delle proteine, monitorata misurandone in continuo l’assorbanza a 280 nm, avviene in presenza di circa 100 mM NaCl nel caso della CRBPI, e di 80 mM nel caso si purifichi la CRBPII. Le frazioni contenenti CRBP vengono identificate, mediante analisi dello spettro d’assorbimento UV-visibile, con le modalità esposte nel caso della cromatografia per gel filtrazione molecolare. Dopo questo passaggio cromatografico la purezza stimata della proteina mediante analisi spettrofotometrica, o elettroforesi in gel denaturante, è del 100%. La quantità di CRBPI e II purificata ad omogeneità mediante questa procedura è di circa 10 mg per litro di coltura batterica.
7.3 Espressione e purificazione di CRABPI ricombinante da Escherichia coli in forma apo e olo
Per poter disporre di CRABPI di ratto, è stato trasformato il ceppo di batteri E.coli XL1B, con il plasmide TT-7 contenente il gene codificante per la proteina, posto sotto il
93

Materiali e Metodi
controllo di un promotore inducibile da IPTG (Isopropil--D-Tiogalattopiranoside), oltre ad un sito che conferisce la resistenza all'ampicillina.Crescita delle colture batteriche ed espressione della CRABPLa precoltura si effettua facendo crescere le cellule batteriche a 37°C in 100 ml di terreno LB (Luria Broth) e 0.1 mg/ml di ampicillina fino a quando queste raggiungono un valore di un valore di OD600 di 0.6-0.7. Circa 6 ml di
cellule batteriche vengono quindi inoculate in 1 l di terreno LB contenente ampicillina (0.1 mg/ml), e incubate a 37 °C. Quando i batteri sono cresciuti fino a raggiungere un valore di OD600 di circa 0.6-0.7 si aggiunge l’induttore IPTG (0.5 mM, stock 100 mM),
che agisce permettendo la trascrizione del gene codificante per la CRABP di ratto da parte della RNA polimerasi del fago T7 presente nel ceppo di E.coli utilizzato. L'induzione avviene per un periodo di 4 ore e 30', al termine del quale le cellule batteriche vengono lavate con tampone TE (50 mM Tris pH 8, 1mM EDTA), e raccolte mediante centrifugazione a 9000g per 10 minuti.Lisi dei batteriTutte le seguenti operazioni, di lisi dei batteri e di purificazione della proteina sono eseguite a 4 °C. I batteri vengono risospesi in 5 volumi di tampone di lisi (50 mM Tris pH 7.9, 15% saccarosio, 1 mM EDTA, 1 mM mercaptoetanolo, 0.05 mM PMSF), e lisati mediante sonicazione. Otto colpi di 15 secondi con pause di circa 1 minuto sono sufficienti per provocare la lisi nella totalità delle cellule. Purificazione della CRABPIL'omogenato viene centrifugato a 30000g per 30', e il surnatante così ottenuto viene dializzato o diluito con tampone di colonna (25 mM potassio fosfato pH 7.2, 200 mM NaCl, 3 mM -Mercaptoetanolo) per essere successivamente caricato su di una colonna a filtrazione molecolare ULTROGEL Aca54, come nel caso della CRBP.Le condizioni di eluizione le attenzioni necessarie per l'utilizzo delle colonne cromatografiche con olo-proteina sono le stesse indicate per la purificazione della CRBP I o II.La olo-CRABPI non è fluorescente, ma, come la olo-CRBP, presenta uno spettro di assorbimento di fluorescenza caratteristico. Questa proprietà viene sfruttata per individuare la presenza della proteina nelle frazioni eluite, mediante analisi dello spettro di assorbimento UV-visibile, che presenta un picco a 350 nm dovuto alla presenza dell’acido retinoico legato. Nel caso sia la apo-CRABP ad essere purificata, questa viene identificata analizzando poi lo spettro di assorbimento di un’aliquota delle frazioni saturate con acido retinoico.A questo punto della purificazione, la purezza della proteina (stimata dall’analisi dello spettro di assorbimento della olo proteina, o mediante elettroforesi) è risultata essere mediamente del 65%.Al fine di rimuovere l'elevata quantità di acidi nucleici presenti nella soluzione proteica eluita dalla colonna di gel filtrazione molecolare, e di purificare ad omogeneità la
94

Materiali e Metodi
proteina, il campione viene applicato su di una colonna a scambio anionico QMA. Nel caso si intenda preparare olo-CRABP, la proteina deve essere saturata prima di questo passaggio cromatografico con un eccesso 1.5 molare di all-trans acido retinico solubilizzato in etanolo. La procedura utilizzata per la purificazione della CRABPI con la colonna di scambio anionico è identica a quella già descritta per la purificazione della CRBP.L’eluizione della proteina, monitorata misurandone in continuo, avviene in presenza una volta raggiunta una concentrazione del gradiente di NaCl pari a circa 120 mM, e le frazioni contenenti CRABP vengono identificate, mediante analisi dello spettro d’assorbimento UV-visibile, con le modalità esposte in precedenza.
7.4 Stima della concentrazione e del grado di saturazione delle retinoid-binding protein
Le proteine leganti i retinoidi presentano, nella forma olo, un caratteristico spettro d’assorbimento UV-visibile con due massimi a 350 e 280 nm; i corrispondenti coefficienti di estinzione molare (indicati in Tabella 26) permettono di quantificare rispettivamente la concentrazione della retinoid-binding protein in forma olo (), e, nota la purezza, quella della proteina totale presente in una soluzione (). Dall’analisi dello spettro è possibile desumere inoltre il grado di purezza e di saturazione della proteina. Infatti il rapporto tra i valori di assorbanza a 350 nm e 280 nm è costante per proteine pure e in forma olo al 100%, e pari a 1.75, 1.65, 1,8 rispettivamente per la CRBPI, II, e la CRABPI (Levin et al., 1990). Nota quindi la percentuale di saturazione, è possibile, misurando il valore di OD350 e di OD280, determinare gli altri parametri relativi alla concentrazione e alla grado di purezza della proteina, e viceversa.
Proteina Peso Molecolare Punto isoelettrico280 nm
(cm-1.M-1)
350 nm
(cm-1.M-1)
CRBP I 14740 4.7 28080 50200
CRBPII 15600 5.6 25506 42700
CRABPI 14700 4.8 27777 50000
Tabella 26: Proprietà fisico-chimiche delle proteine purificate da E.coli
7.5 Elettroforesi di CRBP e CRABP
Al fine di testare la purezza delle retinoid-binding proteins durante, o al termine dei vari passaggi della purificazione, si può effettuare una corsa elettroforetica in condizioni denaturanti o non denaturanti, utilizzando un sistema elettroforetico Mini-protean I della
95

Materiali e Metodi
Bio-Rad. Questo tipo di analisi permette ad esempio in circa 4 ore di risolvere le due forme di CRBPI che sono espresse e purificate da E.coli, che contengono o meno l’amminoacido iniziale metionina (Met+ e Met-). Il gel per effettuare un’elettroforesi analitica in condizioni denaturanti è costituito da poliacrilammide (15% p/v) e da sodiododecilsolfato (SDS, 0.1% p/v). La mancata aggiunta di SDS rende il gel adatto per la separazione delle proteine in condizioni native. Queste condizioni di corsa per il gel, per quanto efficaci, non consentono di risolvere le proteine in forma olo da quelle presenti in forma apo.
7.6 Rimozione del ligando da CRBP e CRABP
Per rimuovere il ligando da olo-CRBP o CRABP, si effettua un’estrazione utilizzando etere etilico. Questa operazione viene eseguita incubando a 4 °C in blanda agitazione, 3 volumi di etere etilico con 1 volume di proteina. Per favorire il processo di rimozione dei retinoidi, è consigliabile sostituire l’etere ogni 40’, monitorando in tempi successivi con misure spettrofotometriche il grado di saturazione residuo della proteina. La rimozione completa del retinolo o dell’acido retinoico dalla CRBPI e CRABPI avviene dopo circa 120 minuti di incubazione, dopodichè occorre eliminare le rimanenti tracce di etere mediante evaporazione sotto corrente d’azoto.Il processo di rimozione del ligando dalle retinoid-binding proteins non porta a denaturazione, nè altera la loro capacità di legare con elevata affinità i retinoidi: infatti, almeno il 95 % delle proteine viene recuperato in seguito ad estrazione, e la CRBPI, così come la CRABPI, sono in grado di legare i retinoidi in modo qualitativamente e quantitativamente identico a quello mostrato prima del trattamento.
7.7 Rimozione dei retinoidi presenti in forma libera
Un metodo rapido ed efficace utilizzato per la rimozione di retinoidi presenti in forma libera (in eccesso o in equilibrio) da soluzioni acquose contenenti CRBP o CRABP, prevede l’utilizzo di microcolonne riutilizzabili contenenti una resina ad elevata lipofilicità (Lipidex 1000) che, grazie alla presenza di catene alifatiche di 15-18 atomi di carbonio quali gruppi funzionali, e una sostituzione in peso pari al 10%, adsorbe specificamente molecole lipidiche quali i retinoidi.La microcolonna è costruita con due un tubi di polipropilene di 1.5 ml, uno, contenente circa 200 µl di resina solubilizzata in etanolo, sul fondo del quale viene praticato manualmente un piccolo foro, e l’altro, contenente il primo tubo, avente la funzione di raccogliere l’eluato. Il campione viene caricato sulla resina, precedentemente lavata con un tampone appropriato, poi raccolto mediante centrifugazione della durata di alcuni secondi. L’eluato viene poi ricaricato due o più volte sulla colonna misurando ogni volta la rimozione dei retinoidi mediante misure spettrofotometriche.
96

Materiali e Metodi
L’efficienza di rimozione per ogni passaggio cromatografico è di circa l’80% del retinolo presente in equilibrio, mentre la rimozione della proteina non è maggiore del 5%. Utilizzando questo tipo di analisi, il ligando complessato con la CRBPI o la CRABPI non viene trattenuto dalla resina, mentre una rimozione di circa il 5% è stata osservata nel caso venga caricata la olo-CRBPII, avente un’affinità per i retinoidi circa 100 volte minore.
7.8 Frazionamento subcellulare
La procedura di frazionamento cellulare per la preparazione del citosol e dei microsomi è stata messa a punto seguendo le indicazioni riportate in letteratura (Posch et al., 1992a).Tutte queste operazioni sono eseguite a 4 °C. L'organo dal quale vengono preparate le frazioni subcellulari, dopo aver eliminato eventuali tonache e membrane esterne, viene lavato con soluzione salina (25 mM Tris pH 7.3, 150 mM NaCl), e frammentato manualmente con un coltello. Si aggiungono poi 2 o 3 volumi di tampone di omogenizzazione (10 mM HEPES pH 7.4 , 0.25 M saccarosio, 2.5 mM -mercaptoetanolo, 0.5 mM DTT, 1 mM leupeptina, 0.5 mM EDTA) per grammo di peso umido nel caso di organi bovini o di ratto, rispettivamente. Nel caso si effettui il frazionamento da colture cellulari, queste sono risospese in un minimo volume di tampone di omogeneizzazione.Per preparare frazioni subcellulari da organi bovini, l’omogeneizzazione avviene in due fasi: il tessuto viene prima trattato con un frullatore Waring Blendor preraffreddato a -20 °C (9 colpi da 10’’, pause di 30’’), quindi con un omogenizzatore vetro/vetro (5 colpi con il tipo A e 2 volte con il tipo B). Per organi o colture cellulari di ratto, la lisi delle cellule è ottenuta esclusivamente utilizzando l’omogenizzatore (vetro/vetro) con le modalità appena descritte.L’omogenato viene quindi frazionato con successive centrifugazioni: la prima a 1400g per 10 minuti; il surnatante ottenuto viene quindi centrifugato a 25000g, o a 15000g nel caso di colture cellulari per 17 minuti. A questo punto il surnatante della precedente centrifugazione viene quindi sottoposto a ultracentrifugazione a 105000g per 1 ora per ottenere così una frazione solubile che verrà utilizzata come citosol, e un pellet che costituisce i microsomi. Questi ultimi sono risospesi in un minimo volume di tampone di omogenizzazione. Al citosol e ai microsomi, è aggiunto KCl ad una concentrazione finale di 150 mM. Le frazioni subcellulari così ottenute vengono suddivise in aliquote che sono immediatamente congelate in azoto liquido e conservate a -80 °C. L’attività di entrambe le frazioni subcellulari si è rivelata essere stabile per almeno 1 anno.La concentrazione proteica è determinata con il metodo del Comassie Blue G-250 (saggio Biorad), utilizzando sieroalbumina bovina come proteina di riferimento.
97

Materiali e Metodi
7.9 Analisi per sedimentazione in gradiente di glicerolo
Il citosol di fegato bovino è stato diluito fino ad una concentrazione proteica di 15 mg/ml e caricato (500 l) su di un gradiente di glicerolo, precedentemente formato, variabile dal 12,5% al 30%. Sono stati utilizzati tre tamponi a differente concentrazione di glicerolo (12,5%, 30%, 50%;) costituiti da 25 mM Tris, pH 7.9, 300 mM ammonio solfato (equivalente a 0.5 M KCl) e 0.2 mM EDTA. Ad ogni tampone è stato aggiunto un mix di inibitori di proteasi tale da ottenere concentrazioni finali di 1 mM DTT; 1 mM Leupeptina; 0.5 mM Benzamidina.Il volume totale dei gradienti è di 9.5 ml: 1 ml costituito dal cuscino di glicerolo al 50%, 8 ml dal gradiente di glicerolo dal 12.5% al 30%, 0.5 ml dalla proteina citosolica e 4 ml dal gradiente. Dopo aver caricato dolcemente il campione, si ultracentrifuga a 287000g per 5 ore e 30 minuti a 0 °C. I gradienti sono quindi frazionati in subaliquote da 500 l, e ciascuna frazione viene saggiata per la sintesi di acido retinoico. Come marker esogeno è stata utilizzata la catalasi a coefficiente di sedimentazione noto le cui bande cadono nelle frazioni 4-5 e 10-11.
7.10 Parziale purificazione dell’attività retinoico biosintetica da citosol di fegato di ratto
Il frazionamento per dimensioni molecolari dell’attività retinoico biosintetica presente nella frazione solubile da fegato di ratto è stato ottenuto mediante cromatografia di gel filtrazione.Circa 5 ml di citosol di fegato di ratto aventi una concentrazione di circa 25 mg/ml sono caricati su di una colonna per filtrazione molecolare Sephacril S-300 HR (2.5 x 90 cm, volume totale 430 ml), precedentemente equilibrata con tampone di colonna (25 mM potassio fosfato pH 7.5, 200 mM NaCl, 3 mM ß-mercaptoetanolo), ed eluiti ad un flusso di 0.35 ml/min raccogliendo frazioni di 4 ml ciascuna. I markers, costituiti da sieroalbumina bovina (P.M.= 67000), alcool deidrogenasi bovina (P.M.= 150000) e da catalasi (P.M.= 230000), sono eluiti dopo 367, 275 e 255 ml rispettivamente.La concentrazione proteica delle frazioni eluite viene misurata mediante il saggio Biorad, o la lettura dell’assorbanza a 280 nm. Le frazioni vengono quindi subaliquotate e congelate a -80 °C.
7.11 Saggio di biosintesi in vitro di Acido Retinoico
Il saggio di reazione per la sintesi di acido retinoico e di retinaldeide viene condotto in tubi di polipropilene da 2.5 ml, in un volume di 200 l.
98

Materiali e Metodi
Il dosaggio avviene in un microambiente normalmente costituito da 150 mM KCl, 50 mM HEPES-K0H, pH 8, 0.05 mM EDTA, 2 mM DTT, 1 µM Leupeptina, in presenza di cofattori (NAD e/o NADP) ad una concentrazione di 2 mM. Condizioni standard di reazione prevedono l’utilizzo di 200g di proteina citosolica e/o 200g di proteina microsomale, e la concentrazione del substrato è di 4M oloCRBPI e/o 4M apoCRBPI.Quando vengono utilizzati come substrato retinoidi in forma libera, questi sono forniti dispersi in etanolo o metanolo in un volume percentuale massimo di 1% (2 µl). I retinoidi sono preparati immediatamente prima della reazione, verificandone spettrofotometricamente la concentrazione e la purezza mediante analisi in HPLC. Condizioni standard per la reazione di biosintesi di acido retinoico prevedono l’incubazione al buio, per 30 minuti, ad una temperatura di 37 °C. La reazione viene terminata mettendo i tubi in ghiaccio, e l’estrazione con solvente organico dei retinoidi viene immediatamente effettuata con le modalità indicate qui di seguito.
7.12 Estrazione dei retinoidi
L’estrazione dei vari retinoidi, siano essi i prodotti di una reazione di biosintesi di acido retinoico o dell’idrolisi dei retinil esteri, oppure ligandi di retinoid-binding proteins, può essere effettuata secondo varie procedure.Il metodo di estrazione seguente, che permette di ottenere una minima isomerizzazione e un buon recupero quantitativo dei vari retinoidi, è utilizzato quando occorre quantificare il prodotto acido retinoico e l’intermedio di reazione retinaldeide del saggio di biosintesi in vitro.La procedura di estrazione dei retinoidi, descritta da Heyman et al. (Heyman et al., 1992), prevede l’aggiunta alla miscela di reazione di 1200 µl di una soluzione di etile acetato e metile acetato (v/v 8:1) a 4 °C, seguita da agitazione per 15”, breve centrifugazione, ed estrazione della fase organica.
Aggiunta Recupero percentuale (%) Forma isomerica di all-trans RA
RA ROH RAld All-trans 9-cis 13-cis
Nessuna 100 42 100 93 2 5
Citosol 72 8 63 93 0.5 6.5
Cit. + Mic. 82 28 70 93 0 7
Tabella 27: Recupero percentuale dei retinoidi estratti con etile acetato e metile acetato. L’estrazione è stata effettuata in condizioni di reazione simulate, utilizzando 200 pmoli di retinoidi in forma all-trans dispersi in 1% etanolo. La quantità di proteina citosolica o microsomale impiegata è di 200 g. L’analisi in HPLC è stata effettuata utilizzando il solvente “D”. Nessuna differenza significativa è stata osservata effettuando l’esperimento con le forme isomeriche 9-cis dei retinoidi.
99

Materiali e Metodi
Questa fase è ripetuta due volte per ottenere il massimo recupero dei retinoidi. In questo caso, i vari isomeri dell’acido retinoico e della retinaldeide, ma non del retinolo, sono estratti con elevata efficienza (Tabella 27), scarsamente isomerizzati, e l’efficienza di estrazione non è influenzata dalla presenza di proteine citosoliche o microsomali.Questo tipo di estrazione, efficace per l’estrazione dei retinoidi in forma libera, non permette una efficiente estrazione di quelli legati alle retinoid-binding proteins. Non può quindi essere utilizzato ad esempio per l’analisi della configurazione isomerica dell’acido retinoico legato alla CRABP.Anche l’estrazione con esano può essere utilizzata per quantificare acido retinoico e retinaldeide; questo tipo di procedura è quella che presenta la minima isomerizzazione dei retinoidi, ma l’efficienza d’estrazione varia notevolmente in presenza di citosol o microsomi (Tabella 28). L’estrazione prevede l’aggiunta alla miscela di reazione 1200 µl di esano a 4°C; si agita per 15”, si centrifuga brevemente, quindi si estrae la fase organica per due volte.
Aggiunta Recupero percentuale (%) Forma isomerica di all-trans RA
RA ROH RAld All-trans 9-cis 13-cis
Nessuna 72 69 78 98 0 2
Citosol 60 35 38 97 1 2
Cit. + Mic. 35 29 28 97 1 2
Tabella 28: Efficienza di recupero di retinoidi estratti con esano. Sono state impiegate le stesse condizioni sperimentali descritte in legenda alla Tabella 27.
Un terzo metodo per l’estrazione dei retinoidi è stato messo a punto da Napoli (Napoli, 1990c). Viene utilizzato per tutti i casi in cui occorre analizzare i vari isomeri del retinolo (es. analisi della configurazione isomerica del retinolo legato alla CRBP), poichè estrae tale molecola con elevata efficienza, senza mostrare una significativa isomerizzazione (Tabella 29). Può essere inoltre utilizzata per l’analisi della configurazione isomerica dell’acido retinoico legato alla CRABP, in quanto estrae questo retinoide con elevata efficienza. L’efficienza di estrazione non dipende dalla quantità di proteine presenti nella miscela di reazione.La procedura prevede l’aggiunta di etanolo a 4 °C contenente 0.2 M HCl e 400 µg/ml BHT (340µl) alla miscela di reazione; si aggiunge esano (1200 µl) a 4 °C, che viene successivamente estratto con le modalità esposte in precedenza ripetendo l’operazione due volte. Questo metodo presenta l’inconveniente, tuttora inspiegato, di presentare, in seguito ad analisi in HPLC, un recupero percentuale di retinaldeide ampiamente superiore al 100%, e non può quindi essere impiegato per il dosaggio di tale molecola.
100

Materiali e Metodi
Aggiunta Recupero percentuale (%) Forma isomerica di all-trans RA
RA ROH RAld All-trans 9-cis 13-cis
Nessuna 100 87 415 95 1 4
Citosol 100 80 322 87 3 10
Cit. + Mic. 96 88 370 85 3 12
Tabella 29: Efficienza di estrazione di retinoidi con esano/ HCl. Sono state impiegate le stesse condizioni sperimentali descritte in legenda alla Tabella 27.
In tutti metodi di estrazione qui descritti, la fase organica contenente i retinoidi viene essiccata sotto corrente di azoto, quindi risospesa in 90 µl della fase mobile per l’immediata analisi in HPLC.
7.13 Analisi in HPLC dei retinoidi
L'analisi dei retinoidi viene eseguita mediante cromatografia ad alta pressione, eluizione isocratica, con un apparecchio Waters 6000A, valvola di iniezione Rheodyne e loop di 600 µl. Il sistema di rilevazione è costituito da un rilevatore di assorbanza Waters 484, ed è posto a 350 nm per l'acido retinoico, 383 nm per la retinaldeide, e 330 nm per il retinolo. Le fasi stazionarie utilizzate sono colonne a fase inversa Novapak C18 (300 mm
x 4.6 mm, Waters), o Hypersyl 5 ODS (250 x 4.6 mm, Shandon). La prima è impiegata nel caso si utilizzino i solventi A, B o D, la seconda invece viene utilizzata con il solvente C.Sono state impiegate quattro differenti fasi mobili per le analisi dei vari retinoidi. Il solvente “A”, costituito da metanolo/acqua (81: 19 v/v), e da 0.01 M ammonio acetato, viene impiegato ad un flusso di 0.7 ml/min per risolvere le varie forme isomeriche di acido retinoico; i tempi di ritenzione per gli isomeri 13-cis, 13/9-cis, 9-cis e all-trans sono rispettivamente di 14’30”, 15’30’’,16’ e 18’ min.Per analizzare la configurazione isomerica dei vari isomeri del retinolo, viene impiegata la fase mobile “B”, (acetonitrile:acqua 70:30 v/v, 0.01M sodio acetato). Ad un flusso di 1.3 ml/min, si osserva l’eluizione degli isomeri del retinolo 13- 9- e all-trans dopo rispettivamente 17 18 e 20 minuti.Il solvente “C”, costituito da acetonitrile/acqua (70: 30 v/v), 0.01 M sodio acetato, è stato utilizzato ad un flusso di 1.5 ml/min per la rilevazione di acido retinoico, retinolo e retinaldeide (tempi di ritenzione 4.5, 18 e 25 min., rispettivamente), senza che avvenga la separazione delle differenti forme isomeriche.L’impiego della fase mobile “D”, messa a punto recentemente, permette l’analisi più rapida ed efficace, in quanto separa in circa 10’ vari isomeri di acido retinoico di retinolo e di retinaldeide, e può così utilizzato per l’analisi del prodotto e dell’intermedio della
101

Materiali e Metodi
reazione di biosintesi di acido retinoico, così come per l’analisi della configurazione isomerica della retinaldeide. Il solvente “D” è costituito da una miscela acetonitrile-acqua 80:20, 0.01 M sodio acetato, acidificato fino a pH 7 con l’aggiunta di HCl; l’eluizione avviene ad un flusso di 1.5 ml/min. I tempi di eluizione per l’acido retinoico 13-cis, 9-cis e all-trans sono 5’50”, 6’, 6’20” minuti rispettivamente; gli isomeri 9-cis/13-cis e all-trans del retinolo sono eluiti dopo 8’ e 8’30” minuti, mentre i tempi di eluizione per gli isomeri 9-cis/13-cis e all-trans della retinaldeide sono di 10’30” e 11’ minuti rispettivamente.Curve di calibrazione per quantificare i differenti isomeri di acido retinoico, retinaldeide e retinolo, sono state costruite misurando l’area e l’altezza dei picchi ottenuti in seguito all’iniezione di quantità note di retinoidi. Si sono verificate condizioni di linearità in tutti i casi esaminati nella costruzione delle curve di calibrazione mediante misura delle aree, mentre nella determinazione dei retinoidi mediante misura dell’altezza dei picchi, si ottengono condizioni di linearità solo in presenza di quantità maggiori di 20 pmoli. Il limite inferiore di rivelazione per i vari solventi utilizzati è di circa 1 pmole.
7.14 Preparazione di 9-cis retinolo mediante riduzione di 9-cis retinaldeide
Il 9-cis retinolo, utilizzato quale substrato per le reazioni di biosintesi di acido retinoico, o quale ligando per testare la formazione del complesso con la CRBP, è stato preparato, non essendo disponibile commercialmente, mediante riduzione di 9-cis retinaldeide.Circa 1 mg di 9-cis retinaldeide viene solubilizzata in 400 µl di metanolo e 100 µl di acqua; si aggiunge quindi l’agente riducente (50 mg di NaB4H4), e si lascia reagire per due minuti. La reazione viene spenta aggiungendo 500 µl di acqua. Si estrae il prodotto della reazione con 2 ml di esano per 3 volte; si essicca in corrente di azoto e si risospende in metanolo. Per ogni preparazione si valuta la purezza isomerica del prodotto ottenuto che è risultato sempre essere almeno al 99% nella forma isomerica 9-cis.
7.15 Misure di fluorescenza
Al fine di studiare l’interazione tra CRBP e retinolo sono state eseguite misure di fluorescenza effettuate utilizzando uno spettrofluorimetro Perkin Elmer LS-50B. E’stata sfruttata la fluorescenza proteica, la cui intensità subisce una sensibile diminuzione a seguito dell’interazione della CRBP con il retinolo. Questa diminuzione è dovuta, molto probabilmente, a trasferimento di energia di eccitazione dai triptofani della proteina al retinolo, che ha un assorbimento nelle regioni di emissione dei triptofani.Il campione è eccitato a 285 nm e l’emissione fluorescente viene rilevata a 340 nm.Ad una soluzione di CRBP 10-7
M in tampone fosfato 25 mM, 0.2 M NaCl, 3 mM -mercaptoetanolo, 2.5 mM benzamidina, sono state aggiunte aliquote di una soluzione di retinolo a diversa concentrazione, solubilizzato in 1% di etanolo. Dopo ogni aggiunta di
102

Materiali e Metodi
retinolo alla soluzione contenente CRBP si è osservata una diminuzione di fluorescenza del triptofano, che si è assunta proporzionale alla quantità di retinolo legato alla proteina (DF parziali). Alle alte concentrazioni di retinolo a cui non si è osservata più alcuna diminuzione di intensità di fluorescenza, si è assunto che i siti della proteina fossero saturati e si è calcolata così il DF totale per la titolazione. Si è assunto infine che i rapporti DF parziale/DF totale per ogni concentrazione di retinolo rappresentassero i valori della saturazione frazionale da cui è stato possibile calcolare Kdiss.
103

Bibliografia
8. BIBLIOGRAFIA
Banaszak, L., Winter, N., Xu, Z., Bernlohr, D.A., Cowan, S. e Jones, T.A. (1994) Advan. Prot. Chem. 45: 89-151
Bavik, C.O., Ericksson, U., Allen, R.A. e Peterson, P.A. (1991) J. Biol. Chem. 266: 14978-14985
Bedo, G., Santisteban, P. e Aranda, A. (1989) Nature 339: 231-234
Ben-Anotz, A., Mokady, S. e Avron, M. (1988) Br. J. Nutr. 59: 443-449
Berni, R., Zanotti, G., Sartori, G. e Monaco H.L. (1993) in “Retinoids: Progress in Research and Clinical Applications” (Livrea, M.A. e Packer, L. Eds.) 7: 91-102 Marcel Dekker, New York
Blaner, W.S., Prystowsky, J.H., Smith, J.E. e Goodman, D.S. (1984) Biochim. Biophys. Acta 794: 419-427
Blaner, W.S., Galdieri, M. and Goodman, D.S. (1987) Biol. Reprod. 36: 130-137
Blaner, W.S., Wei, S., Kurlandsky, S.B. e Episkopou, V. (1994a) in “Retinoids: from Basic Science to Clinical Application” (Livrea M.A. e Vidali, G. Eds.) 53-78, Birkhauser Verlag, Basel
Blaner, W.S. e Olson, J.A. (1994b) in “The Retinoids: Biology, Chemistry, and Medicine” (Sporn, M.B., Roberts, A.B. and Goodman, D.S. Eds.) 229-256 Raven Press Ltd., New York
Blohmoff, R., Holte, K., Naess, L. e Berg, T. (1984) Exp. Cell Res. 150: 186-193
Blohomoff, R., Rasmussen, M., Nilsson, A., Norum, K.R., Berg, T., Blaner, W.S., Kato, M., Mertz, J.R., Goodman, D.S:, Eriksson, U. e Peterson, P.A. (1985) J. Biol. Chem. 260: 13560-13565
Boerman, M.H.E.M. e Napoli, J.L. (1991) J. Biol. Chem. 266: 22273-22278
Boerman, M.H.E.M., e Napoli, J.L. (1995) Biochemistry 34: 7027-7037
Bok, D. e Heller, J (1976) Exp. Eye Res. 22: 395-402
Borden, K.L.B., Boddy, M.N., Lally, J., O’Reilly, N.Y., Martin, S., Howe, K., Solomon, E. e Freemont, P.S. (1995) EMBO J. 14: 1532-1541
Boyd, A.S. (1989) Am. J. Med. 86: 568-574
Breitman, T.R., Selonick, S.E. e Collins, S.J. (1980) Proc. Natl. Acad. Sci. USA 77: 2936-2943
Bychkova V.E., Berni, R., Rossi, G.L., Kutyshenko, V.P. e Ptitsyn, O.B. (1992) Biochemistry 31: 7566-7571
Cavazzini, D., Scita, G., Rossi, G.L., and Ottonello, S. (1994) It. B. Soc. Tran. 1, 305
Cavazzini, D., Galdieri, M. e Ottonello, S. (1996) Biochim. Biophys. Acta, inviato per la pubblicazione
Chai, X., Boerman, M.H.E.M., Zhai, Y. e Napoli, J.L. (1995) J. Biol. Chem. 270: 3900-3904
Chomienne, C. (1994) in “Retinoids: from Basic Science to Clinical Application” (Livrea M.A. e Vidali, G. Eds.) 233-241, Birkhauser Verlag, Basel
Cogan V., Kopelman, M., Mokady, S. e Shinitzky, M. (1976) Eur. J. Biochem. 65: 71-78
Cornic, M., Agadir, A., Degos, L. e Chomienne, C. (1994a), Anticancer Res., 14: 2339-2346
104

Bibliografia
Cornic, M. (1994b) Leukemia 6: 914-917
Cowan, S.W., Newcomer, SE. e Jones, T.A. (1990) Proteins Struct. Funct. Genet. 8: 44-61
Cowan, S.W., Newcomer, S. e Jones, T.A. (1993) J. Mol. Biol. 230: 1225-1246
CreechKraft, J. e Juchau, M.R. (1993) Biochem. Pharmacol. 46: 709-716
Davis, J.T. e Ong, D.E. (1995) Biol. Reprod. 52, 356-364
De Luca, (1991) FASEB J. 5: 2924-2933
Doran, T.I. (1994) in “Retinoids: from Basic Science to Clinical Application” (Livrea M.A. e Vidali, G. Eds.) 315-328, Birkhauser Verlag, Basel
Dowling, J.E. e Wald, G. (1960) Proc. Natl. Acad. Sci. USA 46: 487-499
Duester, G., Shean, M.L., McBride, M.S. e Stewart, M.J. (1991) Mol. Cell. Biol. 11: 1638-1646
Earlye Dimitrovsky, E. 5J. Inv. Med. 43: 337-344
El Akawi, Z. e Napoli, J.L. (1994), Biochemistry 33: 1938-1943
Epstein, J. (1986) Am. Acad. Dermatol. 15: 772-778
Eskild, W., Ree, A.H., Levy, F.O., Jahnsen, T. and Hansson, V. (1991) Biol. Reprod. 44: 55-61
Eskild, W. e Hansson, V. (1994) in "Basic Science and Clinical Aspects of Vitamin A in Health and Disease" (Blomhoff, R., Ed.) 19: 531-559, Marcel Dekker, New York
Fanjui, A., Dawson, M.I., Hobbs, P.D., Jong, L., Cameron, J.F., Harlev, E., Graupner, G., Lu, X., P., e Pfahl, M. (1994) Nature 372: 107-111
Fidge, N.H., Shiratori, T., Ganguly, J. e Goodman, D.S. (1968) J. Lipid Res. 9: 103-109
Fiorella, P.D. e Napoli J.L., (1991a) J. Biol. Chem. 266: 16572-16579
Fiorella, P.D. e Napoli, J.L., (1991b) J. Biol. Chem. 269: 10538-10544
Fiorella, P.D., Giguere, V. e Napoli, J.L. (1993) J. Biol. Chem. 268: 21545-21552
Flower, D.R. (1994) FEBS Letters 354: 7-11
Formelli, F., Clerici, M., Campa, T., Di Mauro, M.G., Magni, A., Mascotti, G., Moglia, D., De Palo, G., Costa, A. e Veronesi, U. (1993) J. Clin. Oncol. 10: 1-8
Fritsch, P. (1992), J. Amer. Acad. Dermatol. 6: S8-S14
Giguere, V., Ong, E.S., Segui, P. e Evans, R.M. (1987) Nature, 330: 624-629
Giguere, V., Lyn, S., e Yip, P. (1990) Proc. Natl. Acad. Sci. USA 87: 6233-6237
Grignani, F., Fagioli, M., Alcalay, M., Longo, L., Pandolfi, P.P, Donti, E., Biondi, A, Lo Coco, F., Grignani, F. e Pelicci, P.G. (1994) Blood 83: 10-15
Gudas, L. (1994), J. Biol. Chem. 269: 22 15399-15402
Harrison, E.H, Blaner, W.S., Goodman, D.S. e Ross, C.A. (1987) J. Lipid Res. 28, 973-981
105

Bibliografia
Harrison, E.H. (1993) Biochim. Biophys. Acta 1170: 99-108
Helgerud P., Petersen, L.B. e Norum, K.R. (1983) J. Clin. Invest. 71: 747-753
Herr, F. e Ong, D.E. (1992) Biochemistry 31: 6748-6755
Heyman, R.A., Mangelsdorf, D.J., Dyck, R.B., Stein, R.B., Eichele, G., Evans, G. e Thaller, C. (1992) Cell 68: 397-406
Hill, D.L. e Grubbs, C.J. (1992) Annu. Rev. Nutr. 12: 161-181
Horlein, J.A., Naar, A.M., Heinzel, T., Torchia, J., Gloss, B., Kurokawa, R., Ryan, A., Kamei, Y, Soderstrom, M., Glass, C. e Rosenfeld, M.G. (1995) Nature 377: 397-404
Huang, H.F.S., Li, M.T., Pogach, L.M. and Qian, L. (1994) Biol. Reprod. 51: 541-550
Jamison, R.S., Kakkad, B., Ebert, D.H., Newcomer, M.E. e Ong, D.E. (1995) Biochemistry 34: 11128-11132
Jörnvall, H., Danilesson, O, Hjelmqvist, L., Hoog, J, Parés, X e Shafqat, J. (1993) in “Enzimology and Molecular Biology of Carbonyl Compounds” (Weiner, H, Crabb, D.W., Flynn, T.G. Eds.) 4: 533-544 Plenum, New York
Kakkad, B.P. e Ong, D.E. (1988) J. Biol. Chem. 263: 12916-12921
Kastner, P. e Chambon, P. (1994) in ”The Retinoids: Biology, Chemistry, and Medicine” (Sporn, M.B., Roberts, A.B. and Goodman, D.S. Eds.) 189-235, Raven Press Ltd., New York
Kato, M., Blaner, W.S., Mertz, J.R., Das, K., Kato, K. e Goodman, D. (1985) J. Biol. Chem. 260: 4832-4840
Kiedel, S., LeMotte, P., e Apfel, C. (1994) Mol. Cell. Biol. 14: 287-298
Kim, K.H. e Griswold, M. (1990) Mol. Endocrinol. 4: 1679-1688
Kim, C.I., Leo, M.A. e Lieber, C.S. (1992) Arch. Biochem. Biophys. 294: 388-393
Kleywegt, G.J., Bergfors, T., Senn, H., Le Motte, P., Gsell, B., Shudo, K. e Jones, T.A. (1994) Structure 2: 1241-1258
Kligman, A.M., Fulton, J.E. e Plewig, G. (1969) Arch. Dermatol. 99: 469-476
Kligman, L.H. (1986) Am. Acad. Dermatol. 15: 779-785
Kurokawa, R., DiRenzo, J. e Boehm, M. (1994) Nature 371, 528-531
Kurokawa, R., Sonderstrom, M., Horlein, A., Halachmi, S., Brown, M., Rosenfeld, M. G. e Glass, C. (1995) Nature 377, 451-454
Labrecque, J., Bhat, P.V. e Lacroix, A. (1993) Biochem. Cell. Biol. 71: 85-89
Labrecque, J., Dumas, F., Lacroix, A. e Bhat, P. (1995) Biochem. J. 305: 681-684
Lampron, C., Rochette-Egly, C., Gorry, P., Dollè, P., Mark, M., Lufkin, T., LeMeur, M. e Chambon, P. (1995) Development 121: 539-548
Lee, M.O., Manthey, C.L., e Sladek, N.E. (1991) Arch. Biochem. Pharmacol. 42: 1279-1285
Lee, M.S., Kliewer, S.A., Provencal, J., Wright, P.E. e Evans, R.M. (1993) Nature 260: 1117-1121
106

Bibliografia
Leid, M., Kastner, P., Durand, B., Krust, A., Leroy, P., Lyons, R., Mendelsohn, C., Nagpal, S., Nakshatri, H., Reibel, C., Saunders, M. e Chambon, P. (1993) Ann. N. Y. Acad. Sci. 684: 19-34
Leo, M.A., Kim, C.I. e Lieber, C.S. (1987) Arch. Biochem. Biophys. 259: 241-247
Levin, A.A., Sturzenbecker, L.J., Kazmer, S., Bosakowski, C., Huselton, G., Allenby, J., Speck, C., Kratzeisen, M., Rosenberg, A., Lovey, A, e Grippo, J.F. (1992) Nature 355: 359-361
Levin, G. e Mokady, S. (1994) Internat. J. Vit. Nutr. Res. 64: 165-169
Levin, M.S., Li, E. e Gordon, J.I. (1990) Methods Enzimol. 189, 506-520
Levin, M.S. (1994) in “Physiology of the Gastrointestinal Tract” (Johnson, L.R. Ed.) 59: 1957-1977, Raven Press Ltd., New York
Li, E., Quian, S., J., Winter, N.S., d’Avignon, A., Levin, M.S. e Gordon, J.I. (1991) J. Biol. Chem. 266: 3622-3629
Lippmann, S.M., Kessler, J.F. e Meyskens, F.L. (1987) Cancer Treat. Rep. 71: 391-405
Lippman, S.M., Heyman, R.A., Kurie, J.M., Benner, S.E. e Hong, W.K. (1995) J. Cell. Biochem. 22: 1-10
Malpeli, G., Folli, C., Cavazzini, D., Sartori, G. e Berni, R. (1996, in corso di pubblicazione) in “Methods in molecular biology: Retinoids” (C. Redfern Ed), The Human Press Inc., Totowa, New Jersey
Mangelsdorf, D.J., Ong, E.S., Dyck, J.A. e Evans, R.M. (1990) Nature 345: 224-229
Mangelsdorf, D.J., Umesono, K e Evans, R.M. (1994) in ”The Retinoids: Biology, Chemistry, and Medicine” (Sporn, M.B., Roberts, A.B. and Goodman, D.S. Eds.) 319-349, Raven Press Ltd., New York
Martin, S., Bradley, S., Cotter, T. Clin. Exp. Immunol. (1990) 79: 448-453
Mc Donald P.N. e Ong, D.E. (1987) J. Biol. Chem. 262: 10550-10556
Mc Donald P.N. e Ong D.E. (1988) J. Biol. Chem. 263: 12478-12482
Miller, W.H., Jakubowski, J.A., Tong, W.P., Miller, V.A., Rigas, J.R., Benedetti, F., Gill, G.M., Truglia, J.A., Ulm, E., Shirley, M. e Warrell, R.P. (1995) Blood 85: 3021-3027
Monaco, H.L., Rizzi, M. e Coda, A. (1995) Science 268: 1039-1041
Muindi, J.R.F. (1994) Leukemia 11: 1807-1812
Nagao, E., Olson, G. (1994) FASEB J. 8: 968-973
Napoli J.L. e Race K.R. (1987) Arch. Biochem. Biophys. 255: 95-101
Napoli, J.L., Pacia, E.B. e Salerno, G.J. (1989) Arch. Biochem. Biophys. 274: 192-199
Napoli, J.L. (1990a) in “Chemistry and Biology of Synthetic Retinoids” (Dawson, M.I., Okamura, W.H. Eds.) 6: 212-229 CRC Press, Boca Raton
Napoli J.L. e Race K.R., (1990b) Biochim. Biophys. Acta 1034: 228-232
Napoli, J. L. (1990c) Methods Enzimol. 189; 470-482
107

Bibliografia
Napoli, J.L., Posch K.C., Fiorella P.D. e Boerman M.H.E.M. (1991) Biomed. Pharmacoter. 45: 131-143
Napoli, J.L., Posch, K.P. e Burns, R.D. (1992) Biochim. Biophys. Acta 1120: 183-186
Napoli, J.L. (1994) in “Basic Science and Clinical Aspects of Vitamin A in Helath and Disease” (Blohomoff, R. Ed.) 6: 135-187 Marcel Dekker, New York
Newcomer, M.E. (1995) FASEB J. 9: 229-239
Noy, N. e Blaner, W (1991) Biochemistry 30: 6380-6386
Ong, D.E. e Chytil, F. (1988) Arch. Biochem. Biophys. 267: 474-478
Ong, D.E., Jamison, R.S. e Newcomer, M.E. (1994) in “Retinoids: from Basic Science to Clinical Application” (Livrea M.A. e Vidali, G. Eds.) 79-89, Birkhauser Verlag, Basel
Ottonello, S., Petrucco, S. e Maraini, G. (1987) J. Biol. Chem. 262: 3975-3981
Ottonello, S., Scita, G., Mantovani, G., Cavazzini, D. e Rossi, G.L. (1993) J. Biol. Chem. 268: 27133-27142
Ottonello S., Cavazzini, D., Scita, G., e Rossi, G.L., (1994) in “Retinoids: from Basic Science to Clinical Application” (Livrea M.A. e Vidali, G. Eds.) 91-102, Birkhauser Verlag, Basel
Petkovich, M., Brand, N.J., Krust, A. e Chambon, P. (1987) Nature 330: 444-450
Porter, S.B., Ong, D.E., Chytil, F. e Orgebin-Christ, M. (1985) J. Androl. 6: 197-208
Posch K.C., Enright, W. e Napoli, J.L. (1989) Arch. Bichem. Biophys. 274: 171-178
Posch, K.C., Boerman, M.H.E.M., Burns, R.D. e Napoli, J.L. (1991) Biochemistry 30: 6224-6230
Posch, K.C., Burns, R.D. e Napoli, J.L. (1992a) J. Biol. Chem. 267: 19676-19682
Posch, K.C. e Napoli, J.L. (1992b) Biochem. Pharm. 43: 2296-2301
Rajan, N., Sung, W.K. e Goodman, D.S. (1990) Biol. Reprod. 43, 835-842
Randolph, R.K. e Ross, C.A. (1991) J. Biol. Chem. 16453-16457; Matsuura, T. e Ross, A.C., (1993) Arch. Biochem. Biophys. 21: 123-130
Rastinejad, F., Perlmann, T., Evans, R. e Sigler, P. (1995) Nature 375: 203-211
Rheaume, E., Snchez, R., Mebarki, F., Gagnon, E., Carel, J.C., Chaussain, J.L., Morel, Y., Labrie, F. e Simard, J. (1995) Biochemistry 34: 2893-2900
Rigtrup, K.M. e Ong, D.E. (1992) Biochemistry 31: 2920-2926
Ross, A.C., (1993) J. Nutr. 123: 346-350
Saari, J.C., Champer, R.J., Batres, M.A., Garwin, G.G., Huang, J., Crab, J.W., e Milam, A.H. (1995) Visual Neuroscience 12: 263-272
Schmitt, M.C. e Ong, D.E. (1993) Biol. Reprod. 49: 972-979
Schule, R., Umesono, K. e Mangelsdorf, D. (1990) Cell 61: 497-504
Schwartz, E.L., Hallam, S., Gallagher, R.C., e Wiernik, P.H. (1995) Biochem. Pharmacol. 50: 923-928
108

Bibliografia
Scott, G.A. e Goldsmith, L.A (1993) J. Inv. Dermatol. 101: 1-8
Shealy, Y.F. (1989) Prev. Med. 18: 624-645
Siegenthaler G., Saurat, J.H. e Ponec, N.M. (1990) Biochem. J. 268: 371-378
Sladek, N.E., Manthey, C.L., Maki, P.L., Zhang, Z. e Lankamer, G.J. (1989) Drug. Metab. Rev. 20: 697-720
Smith, M.A., Parkinson, D.R., Cheson, B. D. e Friedman, M.A.. (1992) J. Clin.Oncol. 5: 839-864
Smith, W., Nakshatri, H., Leroy, P., Rees, J. e Chambon, P. (1991) EMBO J. 10: 2223-2230
Soprano, D.R. e Blaner, W.S. (1994) in “The Retinoids: Biology, Chemistry, and Medicine” (Sporn, M.B., Roberts, A.B. and Goodman, D.S. Eds.) 257-282 Raven Press Ltd., New York
Stahl, W., Sundqvist, A.R., Hanusch, M., Schwarz, W. e Sies, H. (1993) Clin.Chem. 39: 810-814
Stump, D.G., Lloyd, R.S. e Chytil, F. (1991) J. Biol. Chem. 266: 4622-4630
Stunnenberg, H.G. (1993) BioEssays 5: 309-315
Sukov, H., Murakami, K. e Evans, R. (1990) Proc. Natl. Acad. Sci. 87: 5392-5396
Thompson, J.N., McC.Howell J., Pitt G.A., Mc Laughlin, C. (1969) Br. J. Nutr, 23: 471-479
Thompson, J.R., Bratt, J.M. e Banaszak, L.M. (1995) J. Mol. Biol. 252: 433-446
Tzimas, G., Sass, J.O., Ruhl, R., Foerster, M., Klug, S., Collins, M.D. e Nau, H. (1994) in “Retinoids: from Basic Science to Clinical Application” (Livrea M.A. e Vidali, G. Eds.) 179-195, Birkhauser Verlag, Basel
Urbach, J., e Rando, R. (1994a) Biochem. J. 299: 459-465
Urbach, J., e Rando, R. (1994b) FEBS Letters 351: 429-432
Valquist, A., Peterson, P.A. e Wibell, G. (1973) Eur. J. Clin. Invest. 3: 352-363
Vahlquist, A. e Rollman, O. (1987) Dermatologica 175: 20-27
Vasios, G., Gold, J. e Petkovich, M. (1989) Proc. Natl. Acad. Sci. USA 86: 9099-9103
Wald G. (1968), Nature 219: 800-807
Weiss, J.S., Ellis, C.N., Headington, J.T. e Voorhees, J.J. (1988) J. Am. Acad. Dermatol. 19: 169-175
Winter, N., Bratt, J. e Banaszak, L.J. (1993) J. Mol. Biol. 230: 1247-1259
Wolf, G. (1984) Physiol. Rev. 64: 873-937
Yang, Z., N., Davis, G., Hurley, T., Stone, C.L., Li, T.K., e Bosron, W.F. (1994), Alcohol Clin. Exp. Res. 18: 587-591
Yoshida, A., Hsu, L.C. e Davé, V. (1994) Enzyme, 46: 239-244
Yost, R.W., Harrison, E.H. e Ross, C.A. (1988) J. Biol. Chem. 263: 18693-18701
Zanotti, G., Malpeli, G. e Berni, R. (1993) J. Biol. Chem. 268: 24873-24879
Zhuang, J., Liu, Z.P., Jones, T.A., Gierasch, L.M. e Sambrook, J.F. (1992) Proteins, 13: 87-99
109

Bibliografia 110

Pubblicazioni
9. PUBBLICAZIONI
Una parte dei risultati presentati nel presente lavoro di tesi è stato oggetto di recenti pubblicazioni, le quali sono riportate integralmente nelle pagine seguenti:
Ottonello, S., Scita, G., Mantovani, G., Cavazzini, D., and Rossi, G.L. (1993) Retinol Bound to Cellular Retinol-binding Protein Is a substrate for cytosolic retinoic Acid Synthesis.J. Biol. Chem., 268, 36, 27133-27142.
Ottonello, S., Cavazzini, D., Scita, G., and Rossi, G.L. (1994) Retinoic Acid synthesis from Holocellular Retinol Binding Protein. In "Retinoids: from basic science to clinical applications" (Livrea M.A. and Vidali, G. Eds.) 91-102, Birkauser Verlag, Basel.
Malpeli, G., Folli, C., Cavazzini, D., Sartori, G., and Berni, R. (1996) Purification and Fluorescent Tritation of Cellular Retinol-Binding Protein. In “Methods in Molecular Biology: Retinoids” (C. Redfern Ed), The Human Press Inc., Totowa, New JerseyIn stampa
Cavazzini, D., Ottonello, S., and Galdieri, M. (1996) Retinoic Acid synthesis from Holo-Cellular Retinol Binding Protein in the Somatic cells of rat testis.Biochem. Biophys. Res. Commun. In attesa di pubblicazione.
111

RINGRAZIAMENTI
Al termine del dottorato desidero ringraziare: il Professor Gian Luigi Rossi, per i tanti consigli e le discussioni spesso illuminanti, il Professor Simone Ottonello, la cui preziosa e costante guida nella progettazione e nella realizzazione degli esperimenti, nell’analisi e nella valutazione dei risultati, è stata determinante per la riuscita del lavoro. Un grazie anche ai Professori Paolo Colombo e Andrea Mozzarelli per l’attenzione che mi hanno dedicata nel corso degli studi. Ringrazio anche tutti i colleghi con i quali ho collaborato, e in particolare gli amici dell’Istituto di Scienze Biochimiche, la compagnia dei quali ha contribuito a rendere piacevole l’attività di ricerca anche quando questa era più difficile (cioè praticamente sempre).

Ai miei genitori

“Mai dar niente per scontato!”
(Paola Montanari)