Biofisica e Propedeutica Biofisica - Aracne editrice · La Biofisica rappresenta una sintesi...
30
Biofisica e Propedeutica Biofisica Claudio Nicolini
Transcript of Biofisica e Propedeutica Biofisica - Aracne editrice · La Biofisica rappresenta una sintesi...

Biofisicae Propedeutica Biofisica
Claudio Nicolini

Copyright © MMIXARACNE editrice S.r.l.
via Raffaele Garofalo, 133 A/B00173 Roma
(06) 93781065
ISBN 978–88–548–2373–0
I diritti di traduzione, di memorizzazione elettronica,di riproduzione e di adattamento anche parziale,
con qualsiasi mezzo, sono riservati per tutti i Paesi.
Non sono assolutamente consentite le fotocopiesenza il permesso scritto dell’Editore.
I edizione: marzo 2008

V
Indice Prefazione .......................................................................................... XI CAPITOLO I: FISICA E PROPEDEUTICA BIOFISICA ........... 1 1.1. Termodinamica e sue applicazioni ai sistemi biologici ................ 2
1.1.1. Conformazione dei biopolimeri all’equilibrio ........................ 3 1.1.1.1. Denaturazione termica di alti ordini di struttura del DNA 7
1.1.2. Origine ed evoluzione della vita ............................................. 9 1.1.2.1. L’evoluzione chimica .......................................................12 1.1.2.2. L’evoluzione prebiologica................................................18 1.1.2.3. Evoluzione biologica: dagli eobionti alle cellule .............24
1.2. Interazione energia–materia.........................................................29 1.2.1. Radiazione elettromagnetica e regioni spettrali .....................29 1.2.2. L’equazione di Boltzmann .....................................................33
1.2.2.1. Legge di Lambert–Beer....................................................35 1.2.3. Effetti delle radiazioni sulle biostrutture................................37
1.2.3.1. Effetti biologici delle radiazioni non ionizzanti a bassa energia ...........................................................................................38 1.2.3.2. Grandezze utilizzate nella misura delle radiazioni ionizzanti .......................................................................................41
1.3. Meccanica dei fluidi.....................................................................44 1.3.1. Fattori che influenzano lo scorrimento dei fluidi...................44 1.3.2. Elastoviscometro ad alta risoluzione......................................50
1.4. Identificazione di biomolecole e di singoli gruppi atomici .........52 1.4.1. Spettrofotometria in assorbimento .........................................53 1.4.2. Spettri IR di biomolecole .......................................................59 1.4.3. Spettri di fluorescenza............................................................62 1.4.4. Analisi d’attivazione ..............................................................67

VI
1.5. Forma, dimensione e conformazione di biopolimeri................... 71 1.5.1. Elettroforesi e diffusione ....................................................... 72 1.5.2. Sedimentazione...................................................................... 75 1.5.3. Dicroismo circolare ............................................................... 78
CAPITOLO II: INFORMATICA E PROPEDEUTICA BIOFISICA ....................................................................................... 85 2.1. Principi di funzionamento ........................................................... 86 2.2. Memorizzazione delle informazioni............................................ 89 2.3. Linguaggi di programmazione .................................................... 90 2.4. Database ...................................................................................... 92
2.4.1. Protein Data Bank.................................................................. 93 2.5. Prospettive future......................................................................... 94
2.5.1. Sistemi esperti ed intelligenza artificiale in medicina ........... 95 CAPITOLO III: BIOFISICA MOLECOLARE..........................101 3.1. Spettroscopia NMR in soluzione...............................................102
3.1.1. Acquisizione dello spettro NMR .........................................108 3.1.1.1. Chemical shift ................................................................111 3.1.1.2. Accoppiamento spin–spin..............................................114
3.1.2. Spettroscopia NMR bidimensionale di biopolimeri ............116 3.1.2.1. Spettroscopia di correlazione (COSY) ..........................119 3.1.2.2. Spettroscopia Overhauser bidimensionale (NOESY)....121
3.1.3. Applicazione agli spettri su proteine ...................................121 3.1.3.1. Identificazione dei vincoli conformazionali ..................124 3.1.3.2. Calcolo delle strutture 3D..............................................125
3.2. Diffrazione a raggi X di cristalli................................................126 3.2.1. Nucleazione e cristallizzazione di proteine .........................127 3.2.2. La struttura cristallina..........................................................133 3.2.3. Spettri di diffrazione e mappe di densità elettronica ...........137
3.2.3.1. Le sorgenti di raggi X ....................................................141 3.2.3.2. Acquisizione dati cristallografici ...................................145 3.2.3.3. Numero di riflessioni e risoluzione atomica ..................147 3.2.3.4. Densità elettronica e trasformazione di Fourier.............151
3.2.4. Struttura atomica 3D di proteine .........................................155
Indice

VII
3.2.4.1. Metodi per risolvere la fase........................................... 155 3.2.4.2. Le coordinate atomiche ................................................. 157
3.2.5. Sviluppi tecnologici recenti nella cristallografia di proteine....................................................................................................... 158
3.2.5.1. Nanobiocristallografia ................................................... 159 3.2.5.2. Altri tipi di diffrazione .................................................. 164 3.2.5.3. Applicazioni alla medicina............................................ 166
3.3. Scattering in soluzione.............................................................. 170 3.3.1. Via luce polarizzata............................................................. 170 3.3.2. Via neutroni......................................................................... 173 3.3.3. Via luce di sincrotrone ........................................................ 177
CAPITOLO IV: BIOFISICA CELLULARE.............................. 183 4.1. Citometri ................................................................................... 183
4.1.1. Citometri a scansione .......................................................... 184 4.1.1.1. In assorbimento ............................................................. 188 4.1.1.2. In fluorescenza .............................................................. 190 4.1.1.3. In contrasto di fase ........................................................ 193 4.1.1.4. In luce polarizzata ......................................................... 195
4.1.2. Citometri a flusso ................................................................ 196 4.1.2.1. Cell sorter ...................................................................... 200
4.2. Spettrometria di massa di cellule .............................................. 203 4.2.1. Camera di ionizzazione ....................................................... 205
4.2.1.1. Metodi di ionizzazione .................................................. 206 4.2.1.2. Impatto elettronico (EI) ................................................. 207 4.2.1.3. Ionizzazione chimica (CI) ............................................. 208 4.2.1.4. Ionizzazioni per desorbimento ...................................... 209 4.2.1.5. Bombardamento con atomi veloci (FAB) ..................... 209 4.2.1.6. Deassorbimento e ionizzazione laser su matrice (MALDI).................................................................................................... 210 4.2.1.7. Ionizzazione Field Desorption (FD).............................. 211 4.2.1.8. Ionizzazione elettrospray (ESI)..................................... 211
4.2.2. Analizzatore ........................................................................ 213 4.2.2.1. Analizzatore a deflessione magnetica ........................... 215 4.2.2.2. Analizzatori a quadrupolo ............................................. 217 4.2.2.3. Trappola ionica (IT) ...................................................... 218
Indice

VIII
4.2.3. Spettrometro a tempo di volo (TOF) ...................................219 4.2.3.1. Spettrometro a trasformata di Fourier (FTMS)..............220
4.2.4. Rivelatore.............................................................................222 4.2.5. Applicazione allo studio delle cellule..................................224
4.3. Biosensori cellulari ....................................................................226 4.4. La spettroscopia NMR di cellule...............................................232 4.5. Microscopia tunneling e a forza atomica...................................234
4.5.1. STM .....................................................................................235 4.5.2. AFM.....................................................................................239
4.6. Registrazione dei fenomeni elettrici ..........................................241 4.6.1. La tecnica del voltage clamp ...............................................241 4.6.2. Il patch clamp ......................................................................245
CAPITOLO V: BIOFISICA FUNZIONALE SUPRAMOLECOLARE................................................................247 5.1. Unità funzionali biologiche supramolecolari ............................248
5.1.1. Il genoma umano .................................................................249 5.1.1.1 DNA................................................................................251 5.1.1.2. Nucleosoma....................................................................255 5.1.1.3. Fibrosoma ......................................................................257 5.1.1.4. Geni................................................................................260 5.1.1.5. Cromosoma ....................................................................267
5.1.2. Microtubuli e microfilamenti...............................................271 5.1.2.1. Caratteristiche strutturali e funzionali ...........................272
5.1.3. Ribosomi..............................................................................275 5.1.4. Le membrane .......................................................................276
5.1.4.1. Pori della membrana nucleare........................................280 5.1.4.2. Funzione della membrana e fenomeni di trasporto........282 5.1.4.3. Potenziale elettrico e selettività ionica della membrana 283
5.2. Funzioni cellulari e meccanismi molecolari..............................288 5.2.1. Trasformazione neoplastica, proliferazione ed invecchiamento.......................................................................................................288
5.2.1.1. Trasformazione neoplastica ...........................................288 5.2.1.2. Proliferazione cellulare ..................................................290 5.2.1.3. Invecchiamento cellulare ...............................................296 5.2.1.4. Meccanismi molecolari e cellulari di controllo .............298
Indice

IX
5.2.1.4.1. Superelicità negativa ............................................... 298 5.2.1.4.2. Accoppiamento nucleo–cellula ............................... 300 5.2.1.4.3. Ruolo dell’acqua...................................................... 302
5.2.2. Eccitazione nervosa............................................................. 306 5.2.2.1. Il potenziale di membrana a riposo ............................... 308 5.2.2.2. La propagazione del potenziale d’azione ...................... 312 5.2.2.3. Le equazioni di Hodgkin e Huxley ............................... 315
CAPITOLO VI: BIOFISICA MEDICA...................................... 319 6.1. Diagnostica Medica Biofisica ................................................... 320
6.1.1. Tomografia .......................................................................... 321 6.1.1.1. Tomografia assiale computerizzata a raggi X (TAC) ... 326 6.1.1.2. Tomografia ad Emissione di Positroni (PET) ............... 330
6.1.1.2.1. Tomografia ad emissione di positroni a tempo di volo (TOF)....................................................................................... 336 6.1.1.2.2. Applicazioni cliniche della tomografia ad emissione................................................................................................. 336
6.1.1.3. Tomografia a Risonanza Magnetica Nucleare (TRMN)341 6.1.1.3.1. Tempi di rilassamento nelle coordinate rotanti ....... 355 6.1.1.3.2. Misura dei tempi di rilassamento ............................ 358 6.1.1.3.3. Applicazioni in medicina......................................... 361
6.1.2. Spettroscopia NMR in vivo................................................. 368 6.1.2.1. Applicazioni agli organi umani ..................................... 373
6.1.3. Laser.................................................................................... 378 6.1.3.1. Principi di funzionamento ............................................. 380
6.1.3.1.1. Sorgenti laser ........................................................... 382 6.1.3.2. Applicazioni in campo medico...................................... 383
6.1.4. Microarray........................................................................... 385 6.1.4.1. Genomica....................................................................... 387 6.1.4.2. Proteomica..................................................................... 390
6.2. Biofisica degli organi di senso.................................................. 395 6.2.1. Traduzione sensoriale ......................................................... 396 6.2.2. Risposte elettriche ............................................................... 397 6.2.3. Codifica dei segnali............................................................. 399 6.2.4. Modelli ................................................................................ 402 6.2.5. Visione ................................................................................ 403
Indice

X
6.2.5.1. Fotorecettori...................................................................406 6.2.5.2. Le componenti retiniche ................................................417 6.2.5.3. Sensibilità assoluta e adattamento .................................420
6.2.6. Udito ....................................................................................422 6.2.6.1. Principi fisici..................................................................423 6.2.6.2. Struttura e funzione dell’orecchio..................................426
6.3. Olfatto........................................................................................431 6.3.1. Trasduzione chemo–olfattiva...............................................435
Appendici Abbreviazioni ...................................................................................441 Unità di misura e costanti fisiche .....................................................443 Indice analitico .................................................................................445
Indice

XI
Prefazione
Fra gli elementi innovativi del panorama scientifico internazionale
vi sono sicuramente gli argomenti qui raggruppati come Biofisica e Propedeutica Biofisica, di particolare rilevanza per gli studi di Medicina e per l’Alta Formazione Biomedica (Master e Dottorati di Ricerca). La Biofisica rappresenta una sintesi degli aspetti avanzati e quantitativi dello studio della materia vivente dai suoi elementari costituenti molecolari, i biopolimeri, sino all'uomo, nonché delle sue applicazioni pratiche e tecnologiche all’intersezione fra le scienze fisiche e le scienze della vita.
Molti aspetti della Fisica e dell’Informatica sono necessariamente propedeutici alla Biofisica e formano quindi il cuore dei primi due Capitoli, mentre i restanti quattro Capitoli forniscono il necessario bagaglio quantitativo, strutturale e strumentale a livello molecolare, supramolecolare, cellulare e medico per affrontare la costante evoluzione scientifica, tecnologica e professionale della BioMedicina.
Il libro risulta anche dalla preziosa collaborazione dei seguenti assistenti senior della Cattedra di Biofisica, tutti dotati di Dottorato di Ricerca in Biofisica, co–estensori dei paragrafi del Libro indicati fra parentesi: Eugenia Pechkova (3.2.1, 3.2.2, 3.2.3, 3.2.4, 3.3.3); Rosanna Spera (4.4.1, 4.4.2, 4.4.3); Enrico Stura (2.1, 2.2, 2.3, 4.3, 6.4.1); Francesca Vasile (3.1.1, 3.1.2, 3.1.3).


1
CAPITOLO I FISICA E PROPEDEUTICA BIOFISICA
Il ciclo vitale di una cellula e l’organizzazione dei biopolimeri in strutture ben definite sono tipici processi termodinamici, poiché en-trambi hanno inizio con la “cattura” di fotoni dal Sole e procedono attraverso una serie di scambi di energia con l’ambiente (Figura 1.1).
Figura 1.1. Esemplificazione di processi biologici che iniziano con la cattura di fo-toni dal Sole e procedono attraverso scambi d’energia con l’ambiente.

Capitolo I
2
1.1. Termodinamica e sue applicazioni ai sistemi biologici
Gli approcci teorici a tali processi, sono essenzialmente riconduci-bili in due ambiti termodinamici, drasticamente diversi tra loro ma equivalenti nella capacità di spiegazione.
Termodinamica classica, in cui ogni sistema di n mole è definito in termini di parametri macroscopici, quali pressione (P), volume (V) e temperatura assoluta (T), tra loro collegati attraverso la legge dei gas ideali di Boyle e Charles:
PV=nRT
ove R è la costante universale dei gas (8,31 × 107 erg/Kmol). Ogni mole contiene lo stesso numero di molecole NA (dalla legge d’Avogadro, NA = 6023 × 1020 molecole/mol per qualsiasi gas o so-stanza chimica).
Meccanica statistica in cui ogni sistema macromolecolare è defini-to in termini di parametri microscopici, quali posizione (x, y, z) e quantità di moto (Px, Py, Pz) di ognuna delle sue N molecole (N=nNA). Sono quindi necessarie 6N coordinate per caratterizzare ogni sistema macromolecolare al livello molecolare. Per ogni proprietà macrosco-pica, M, la media di tutti i suoi possibili valori, assunti all’equilibrio dal sistema entro il volume V, è determinata dalla densità di probabili-tà:
∫∫= dVMdVMV
/
Le leggi fondamentali e le funzioni di stato più importanti, quali entalpia (H) entropia (S) ed energia libera di Gibbs (G), eguale a
STHG ∆−∆=∆ , sono riassumibili nell'ambito dei due approcci di-stinti e direttamente correlati della termodinamica classica e della meccanica statistica; gli stessi principi fondamentali vengono quindi applicati allo studio delle strutture biologiche ed alla determinazione della direzione e condizione di equilibrio delle molteplici reazioni chimiche aventi luogo nelle cellule viventi e/o in soluzioni di biopo-limeri. Ulteriori dettagli possono essere rinvenuti in classici testi di riferimento ed in particolare in Nicolini e Rigo, Biofisica e Tecnologie Biomediche, Zanichelli Editore, 1992 (ora fuori stampa).

Fisica e Propedeutica Biofisica
3
I fenomeni irreversibili tipici dei processi di trasporto occorrenti in natura nonché l’origine e l’evoluzione della vita, sono due applicazio-ni concrete della termodinamica, utili alla comprensione sia delle for-ze e delle strutture in gioco, sia della loro mutua interazione nel de-terminare il progredire ed il funzionamento dei sistemi biologici.
1.1.1. Conformazione dei biopolimeri all’equilibrio
Una domanda che frequentemente insorge è come le cellule possa-no esibire un tale comportamento ordinato nonostante la presenza di un forte elemento di disordine provocato dall’incessante agitazione termica dei singoli atomi. Questo paragrafo tenterà di dare una rispo-sta a questa domanda nell’ambito della termodinamica e della mecca-nica statistica.
Si consideri una molecola di DNA a doppia elica (Figura 1.2) che in condizioni native è nota assumere un’unica struttura secondaria ca-ratterizzata da una sorta di conformazione a spago (la probabilità ter-modinamica dello stato nativo ΩN è 1).
Figura 1.2. Rappresentazione schematica degli stati energetici di una proteina, nell’ipotesi che la stessa assuma una sola conformazione nello stato nativo e più conformazioni nello stato denaturato.
In condizioni di denaturazione termica il DNA subisce un drastico cambiamento strutturale dando origine a due singole eliche caratteriz-zate da una sorta d’aggomitolamento a caso, in cui ogni nucleotide può ruotare liberamente intorno ai legami della catena. Quest’avvolgimento a caso può assumere N conformazioni fortemente influenzate dal solvente ed è caratterizzato quindi dall’assenza di

Capitolo I
4
un’unica struttura tridimensionale (cioè, la probabilità termodinamica dello stato denaturato è ΩD = N).
Dall’equazione di Boltzmann, il rapporto fra il numero di molecole di DNA nello stato nativo (nN) verso quella nella forma denaturata (nD) è
[ ] [ ][ ]RTggRT
N
D
N
D NDNDND eegg
nn /))/ln(/)( εεεε −−−− ==
oppure
[ ] [ ]kTSTHkTkT
N
D eenn
NDND /)(/)/ln()( ∆−∆−ΩΩ−−− == εε
assumendo che la degenerazione per livello di energia sia uguale alla probabilità termodinamica (gN = ΩN e gD = ΩD). Va ricordato che la costante di Boltzmann k viene espressa in erg/grado × molecola, ed è collegata alla costante universale dei gas R dalla relazione ANRk /= , ove NA è il numero di Avogadro. Abbiamo inoltre assunto che ∆U < = εD – εN) ≅ ∆H. In unità molari, possiamo infine scrivere
[ ])/( RTG
N
D o
enn ∆−=
dove Keq è la costante di equilibrio. Tale relazione fa vedere come essendo l’energia libera di denatu-
razione, sempre negativa (∆G = RT1 o gKeq < 0), il senso spontaneo della reazione sarebbe sempre verso lo stato denaturato. Di conse-guenza, senza la presenza di certi tipi di legame (quali i legami idro-geno che infatti esistono in questo caso), la doppia elica, che è la struttura tipica del DNA nello stato nativo, non potrebbe essere man-tenuta.
Questa considerazione si applica alla maggior parte dei biopolime-ri, le cui strutture secondarie e di ordine più alto sono mantenute e modulate da legami deboli tipo quello idrogeno, oltre che dall’interazione soluto–solvente. Infatti, l’autoassemblaggio di nume-rosi importanti biomolecole (quali microtubuli, fosfolipidi e collage-

Fisica e Propedeutica Biofisica
5
ne) avviene in parte a seguito delle loro interazioni con solventi polari (principalmente l’acqua) che dà origine a legami di natura idrofobica, ossia ad una tendenza dei gruppi non polari a aggregarsi in presenza dell’acqua.
Paradossalmente in questi casi la stabilità della conformazione rag-giunta all’equilibrio aumenta all’aumentare della temperatura (Figura 1.3); esattamente il contrario di quanto precedentemente mostrato per DNA e proteine.
Figura 1.3. (a) Autoassemblaggio (T = 37°C) e disassemblaggio (T = 10°C) del virus RNA del tabacco. (b) Autoassemblaggio (T > 37°C) e disassemblaggio (T < 0 °C) di un microtubulo da monomeri di tubulina α e ß. (e) Autoassemblaggio di liposomi da lipidi anflipatici secchi in acqua.
Per evitare l’interazione con i gruppi non polari presenti in tali bio-polimeri le molecole d’acqua aumentano il loro grado di disordine e quindi la loro entropia (∆SA>0). Questo spiega perché l’autoassemblaggio di tali biopolimeri dalle loro sottounità (monome-ri), che è associato ad un aumento del grado di ordine e quindi ad una diminuzione dell’entropia (∆SP<0), avviene spontaneamente pur es-

Capitolo I
6
sendo di natura endotermica la relativa denaturazione 0>∆ BH . Que-sto consegue dal fatto che l’energia libera totale, data dall’equazione
( )ABB SSTHG ∆+∆−∆=∆ , è negativa per il contributo entropico po-sitivo (ed in eccesso rispetto al contributo negativo del biopolimero) delle molecole di acqua, le quali, durante il processo di aggregazione, diminuiscono le interazioni con le singole biomolecole a cui erano inizialmente legate. In altre parole, l’autoassemblaggio di tali biopo-limeri la cui stabilità aumenta all’aumentare della temperatura (più grande è T più negativo diventa ∆G), è guidato dalla presenza di un termine entropico legato alle interazioni idrofobiche tra le sottounità che le compongono. In assenza di tali interazioni, ∆G sarebbe sempre maggiore di zero e quindi la formazione di tali biopolimeri non po-trebbe mai avvenire spontaneamente a qualsiasi temperatura.
Classici esempi di tale fenomeno sono: 1) il virus RNA del tabacco che autoassembla da 2100 sottounità
proteiche (di circa 17000 dalton l’una); stabile a T > 16°C e depolimerizza a T < 16°C (Figura 1.3.a);
2) i microtubuli, che si autoassemblano in modo irreversibile da monomeri di tubolina α e ß; stabile a ≥, 37 °C ed instabile de-polimerizzandosi a ≤ 15°C (Figura 1.3.b);
3) i liposomi, che si formano per addizione di lipidi anfipatici a soluzioni acquose (Figura 1.3.c);
4) la formazione spontanea in modo irreversibile di collagene da polipeptidi.
In soluzione, i biopolimeri possono assumere varie conformazioni riconducibili a tre grosse classi. 1) A matassa (random coil), per cui non vi è un’unica struttura tridi-mensionale ed il biopolimero è costantemente modificato dall’interazione del solvente; in questo caso, si parla soltanto di di-mensioni medie quali il raggio di girazione RG il cui quadrato è dato da:
∑= NRR iG /22
ove Ri è la distanza media dei singoli segmenti dal centro di massa ed N è il numero dei segmenti che compongono il biopolimero.

Fisica e Propedeutica Biofisica
7
Catene di biopolimeri lineari (DNA a singola elica), libere di ruota-re intorno ai legami e caratterizzate da modeste interazioni fra gruppi laterali, tendono ad assumere tali conformazioni, analisi statistiche indicano la presenza in media di una simmetria sferica intorno al cen-tro di massa di ogni catena ed un raggio di girazione proporzionale alla radice quadrata del numero dei segmenti che la compongono (ad es., nel caso del DNA al numero di basi fra loro distanti circa 3,4 Å). 2) A corda (rodlike) per cui vi è un’unica struttura tridimensionale tipicamente a forma di elica. Per tali biopolimeri, quali polinucleotidi a doppia elica e polipeptidi ad α–elica, le proprietà in soluzione sono dominate dalla lunghezza della corda e non tanto dal suo diametro. 3) Globulare, per cui vi sono interazioni tra gruppi laterali che tendo-no ad aggomitolare le catene in un’impacchettatura a conformazione globulare.
Un classico esempio sono le proteine globulari, che assumono una forma elicoidale–prolata (del tipo a sigaro) o oblata.
1.1.1.1. Denaturazione termica di alti ordini di struttura del DNA
Le transizioni termiche di macromolecole sia semplici (quali il DNA nel suo basso ordine di struttura secondaria precedentemente descritta) che complesse (quali la cromatina negli alti ordini di struttu-ra del DNA) sono quantificabili attraverso la misura della loro iper-cromicità, ossia dell’aumento in assorbimento ottico che accompagna sia la transizione superelica → elica (da un alto ordine di struttura a superelica ad una struttura secondaria ad elica) sia a quella d’ordine inferiore helix–coil.
Comunque, è solo attraverso misure di dicroismo circolare che è possibile discriminare tra tali transizioni, rivelando anche i più sottili cambiamenti nell’asimmetria della superelica (caratterizzati da un aumento del segnale di dicroismo circolare) ben prima della denatu-razione della doppia elica di DNA (caratterizzata da una diminuzione del segnale di dicroismo circolare come visto successivamente).
Appare evidente come la stabilità della doppia elica, in particolare la sua temperatura di fusione T, aumenti significativamente nella cro-matina per l’interazione con le proteine istoniche cromosomiche ad essa legate; tale determinazione, unitamente a quella della temperatura

Capitolo I
8
di fusione della superelica, costituiscono ulteriori esempi del tipo di importanti informazioni che si possono trarre da semplici esperimenti di denaturazione termica.
Tipicamente, tali misure ottiche (dicroismo circolare ed assorbi-mento ottico) sono compiute in soluzioni tamponate contenenti il bio-polimero, in funzione della temperatura, attraverso spettropolarimetri e spettrofotometri che utilizzano speciali celle di quarzo fuso, in cui viene fatta circolare opportunamente l’acqua per consentire il control-lo della temperatura. Durante tali studi di denaturazione termica ven-gono mantenute fisse le lunghezze d’onda, in corrispondenza alla banda di assorbimento del cromoforo di interesse (ad esempio, 260 nm per gli acidi nucleici e 220 nm per le proteine).
Una stima dell’ammontare e del tipo di legami che mantengono le strutture in esame si ha dalla determinazione con microcalorimetri a scansione della loro capacità termica a pressione costante (variazione d’entalpia) in funzione della temperatura.
Figura 1.4. Capacità termica (cal/g) contro temperatura per le stesse soluzioni di DNA (a) e cromatina nativa (b) di Dna in 1 mM Tris-HCl, pH 8.
Tali misure, condotte con microcalorimetri differenziali a scansio-ne, sono esemplificate in Figura 1.4 per le stesse soluzioni di DNA e cromatina di cui sopra. Esse consentono di determinare univocamente il numero di transizioni presenti in un vasto intervallo di temperature (ben oltre la temperatura di ebollizione dell’acqua, in cui sono invece confinate le analisi spettrofotometriche e spettropolarimetriche di cui

Fisica e Propedeutica Biofisica
9
sopra); nel caso di un biopolimero complesso ed eterogeneo quale la cromatina se ne identificano ben tre (Figura 1.4).
Tabella 1.1. Dati calorimetrici relativi alle transizioni conformazionali di alcuni im-portanti biopolimeri.
Biopolimero Reazione pH Solvente Temperatura di denaturazione
(°C)
∆H (kcal)
Ribonucleasi A Denaturazione 2,8 0,15 m KCl 43 70/mol Mioglobina Denaturazione 4,5 0,15 m KCl 30 40/mol Fibrina Polimerizza-
zione 6,08 1,0 m NaBr 25 -19/mol
Poli–L–glutammato
Denaturazione 5 0,1 m LiCl 30 -1,1/mol resi-duo
DNA di salmone Denaturazione 6 0,1 m NaCl 25 8,3/mol coppia di basi
Tripsina Denaturazione 2 0,1 m NaCl 25 8,0/mol Poli A (doppia elica)
Denaturazione 4,1 0,1 m K Cl 25 2,7/mol nucle-otide
DNA di riccio marino
Denaturazione 6 0,1 m NaCl 25 8,1/mol coppia di basi
La Tabella 1.1 riepiloga i dati calorimetrici di transizione confor-mazionali di alcuni importanti biopolimeri, in particolare la variazione di entalpia legata al calore assorbito nella transizione alla struttura primaria.
1.1.2. Origine ed evoluzione della vita
L’origine della vita e la sua evoluzione sono fenomeni cosmici, che coinvolgono tanto l’Universo fisico quanto la materia vivente. Gli a-spetti che essi assumono in scenari cosi diversi come la prima sintesi di nuclei atomici e lo sviluppo della corteccia cerebrale, differiscono fra loro al punto da renderne difficilmente discernibili i lineamenti comuni che pure esistono, come cercheremo di dimostrare. L’intuizione dell’esistenza di un coordinamento globale fra fenomeni non collegati od apparentemente contraddittori è una conquista recen-te. Nel campo biologico, dove sono apparse prima, le teorie evoluzio-nistiche nascono alla metà del secolo diciannovesimo con J. B. de Lamarck e C. R. Darwin. Più recente è la constatazione dell’esistenza di fenomeni evolutivi dell’universo fisico, su scale di spazio e di tem-po così estese, che solo ultimamente si è riusciti ad ottenerne una co-

Capitolo I
10
noscenza operativa. Appena trent’anni fa una delle più note teorie co-smologiche postulava che l’universo si trovasse in uno stato staziona-rio. Una percezione cosi tardiva dell’intrinseca direzionalità dei pro-cessi è dovuta all’impossibilità di osservare l’evoluzione su scala ri-dotta. Degli astri muoiono ed al loro posto altri se ne formano, cosic-ché si può avere l’impressione che globalmente l’universo rimanga immutato. L’espansione cosmica, che è il segnale più facilmente per-cepibile di un cambiamento nel corso del tempo, è osservabile solo a livello intergalattico. La graduale variazione della composizione ele-mentare della materia nell’universo, dovuta alla nucleosintesi nelle stelle, cambia le popolazioni stellari su una scala di tempo così grande che solo recentemente se n’è avuta verifica, in gran parte indiretta. L’esistenza di uno stato singolare dello spazio–tempo all’origine del nostro Universo fisico è nozione del tutto induttiva e recentissima, an-che se consolidata. La nozione di un Universo in trasformazione ha potuto venir acquisita solo quando l’uomo è riuscito ad estendere le sue capacità di osservazione su scale spazio–temporali smisurate.
Non altrimenti nell’evoluzione delle specie viventi: i singoli indi-vidui nascono e muoiono, conservando per lo più le caratteristiche proprie della specie. L’osservazione, già di per sé difficile, del lentis-simo processo di mutazione ed adattamento sembra perdere gran parte della sua significatività di fronte alla constatazione che alla fine anche le specie si estinguono. In effetti, si stima che i sei o sette milioni di specie viventi note si vadano estinguendo al ritmo di diecimila per an-no venendo via–via rimpiazzate da un numero all’incirca equivalente di altre di nuova formazione, non necessariamente più perfezionate delle precedenti. Per riuscire a percepire delle tendenze d’insieme di questo continuo rimescolio, occorre estendere il campo d’osservazione fino ad includervi l’intera biosfera e considerare con-grui intervalli temporali, valutabili in centinaia di milioni, o miliardi di anni. È probabile che l’esempio più convincente dell’esistenza di una direzionalità intrinseca negli eventi e del coordinamento fra fe-nomeni distinti venga dalle circostanze che hanno portato all’apparizione dei primi organismi viventi sulla Terra. Le tappe prin-cipali percorse possono oggi essere identificate con relativa sicurezza e, cosa più importante, è possibile individuare il tipo di forze che han-

Fisica e Propedeutica Biofisica
11
no operato, nonché i rapporti fra loro e le strutture entro cui esplica-vano la propria azione. Cerchiamo in primo luogo di definire e circo-scrivere il campo che vogliamo trattare entro il più generale quadro dell’evoluzione cosmica (Figura 1.5), ove rientra nel breve intervallo ombreggiato alla estrema destra dello schema in figura.
Figura 1.5. Principali stadi evolutivi del cosmo e della biosfera. Sulle ascisse i tem-pi, in secondi, sulle ordinate le temperature medie dell’Universo ovvero le densità medie dell’energia. Esistono varie ragioni per ritenere che la funzione “produzione di entropia” P diminuisce proporzionalmente alla densità di energia media. Le ordi-nate potrebbero quindi essere alternativamente rappresentate in funzione dell’integrale della somma di tutti i prodotti fra le forze generalizzate ed i flussi da queste generati, nell’intero Universo.
Il periodo di tempo coperto è dell’ordine del miliardo di anni ed il luogo la superficie della Terra. Possiamo suddividere il campo in due parti: l’evoluzione chimica e quella più propriamente prebiologica. Soffermiamoci ancora per un momento ad osservare la Figura 1.5 nel suo complesso: sulle ordinate sono riportate le temperature o, ciò che è lo stesso, le densità di energia. Questa scala potrebbe forse essere

Capitolo I
12
espressa anche in termini della rapidità di produzione d’entropia P nell’Universo. Tale grandezza può venire rappresentata come l’integrale dei prodotti delle forze generalizzate X e dei flussi J aventi luogo nel sistema:
∫∑=V ki
kiki dVXJP,
,, )(
V essendo nel nostro caso il volume dell’Universo. In questi termini la figura suggerisce, sulla base del teorema di Glansdorff–Prigogine, che la diminuzione di P nel tempo possa indicare l’avvio dell’Universo ad uno stato stazionario, ma ovviamente l’argomento è controverso.
1.1.2.1. L’evoluzione chimica
L’Universo che emerge dalla prima nucleosintesi dopo il disaccop-piamento dei neutrini dalla materia (Figura 1.5) è costituito quasi e-sclusivamente di idrogeno ed elio, con l’aggiunta di litio, berillio e boro in concentrazioni estremamente ridotte. Le condizioni dell’espansione iniziale (big bang) hanno predeterminato le linee ge-nerali degli eventi successivi, in termini della loro compatibilità con le condizioni al contorno nel cui ambito si devono svolgere. L’evoluzione è già insita nelle regole del gioco — i valori delle co-stanti fondamentali, h, costante di Planck; C, velocità della luce; G, costante gravitazionale ecc. — e nella natura dei mattoni fondamentali — quarks, leptoni e fotoni — in un modo non ancora chiaro e che co-munque portò alle disomogeneità nella distribuzione della materia.
Queste disomogeneità consentirono alla forza di gravità di prevale-re localmente, agglomerando le grandi strutture dell’Universo — i su-perammassi e le galassie — e provocando al loro interno la condensa-zione di stelle. In molti di questi astri la temperatura aumentava con-temporaneamente all’addensarsi della materia fino a produrre condi-zioni adatte alla sintesi di nuclei complessi. L’intrinseca instabilità degli edifici stellari ne causava prima o poi l’esplosione, durante la quale parte del materiale raggiungeva condizioni in cui poteva com-piersi la seconda nucleosintesi producendo tutti i nuclei stabili oltre a molti altri instabili. L’esplosione dell’astro disperdeva il materiale ne-osintetizzato nello spazio circostante dove, prima o poi, entrava a far
Fisica e Propedeutica Biofisica
13
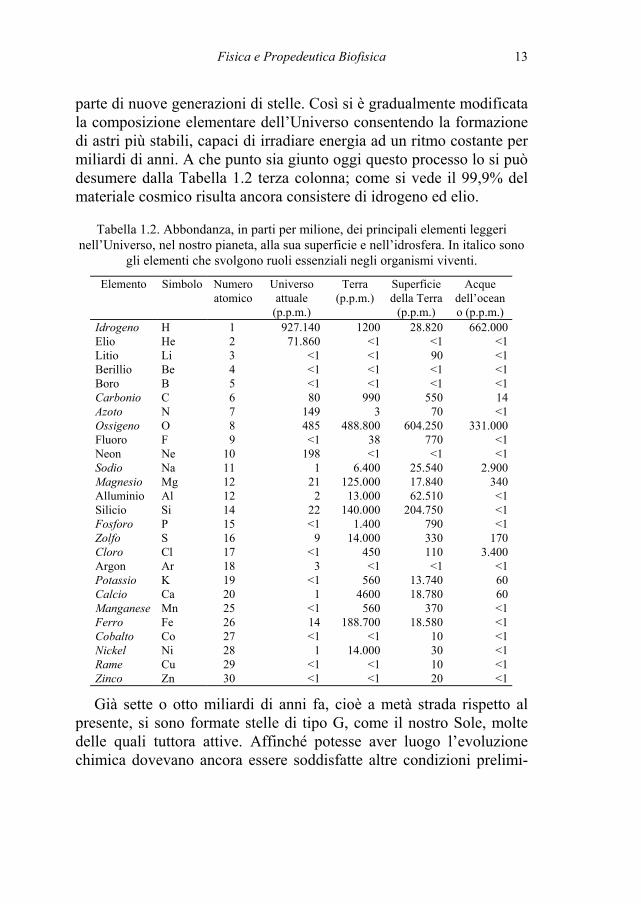
parte di nuove generazioni di stelle. Così si è gradualmente modificata la composizione elementare dell’Universo consentendo la formazione di astri più stabili, capaci di irradiare energia ad un ritmo costante per miliardi di anni. A che punto sia giunto oggi questo processo lo si può desumere dalla Tabella 1.2 terza colonna; come si vede il 99,9% del materiale cosmico risulta ancora consistere di idrogeno ed elio.
Tabella 1.2. Abbondanza, in parti per milione, dei principali elementi leggeri nell’Universo, nel nostro pianeta, alla sua superficie e nell’idrosfera. In italico sono
gli elementi che svolgono ruoli essenziali negli organismi viventi.
Elemento Simbolo Numero atomico
Universo attuale
(p.p.m.)
Terra (p.p.m.)
Superficie della Terra
(p.p.m.)
Acque dell’oceano (p.p.m.)
Idrogeno H 1 927.140 1200 28.820 662.000 Elio He 2 71.860 <1 <1 <1 Litio Li 3 <1 <1 90 <1 Berillio Be 4 <1 <1 <1 <1 Boro B 5 <1 <1 <1 <1 Carbonio C 6 80 990 550 14 Azoto N 7 149 3 70 <1 Ossigeno O 8 485 488.800 604.250 331.000 Fluoro F 9 <1 38 770 <1 Neon Ne 10 198 <1 <1 <1 Sodio Na 11 1 6.400 25.540 2.900 Magnesio Mg 12 21 125.000 17.840 340 Alluminio Al 12 2 13.000 62.510 <1 Silicio Si 14 22 140.000 204.750 <1 Fosforo P 15 <1 1.400 790 <1 Zolfo S 16 9 14.000 330 170 Cloro Cl 17 <1 450 110 3.400 Argon Ar 18 3 <1 <1 <1 Potassio K 19 <1 560 13.740 60 Calcio Ca 20 1 4600 18.780 60 Manganese Mn 25 <1 560 370 <1 Ferro Fe 26 14 188.700 18.580 <1 Cobalto Co 27 <1 <1 10 <1 Nickel Ni 28 1 14.000 30 <1 Rame Cu 29 <1 <1 10 <1 Zinco Zn 30 <1 <1 20 <1
Già sette o otto miliardi di anni fa, cioè a metà strada rispetto al presente, si sono formate stelle di tipo G, come il nostro Sole, molte delle quali tuttora attive. Affinché potesse aver luogo l’evoluzione chimica dovevano ancora essere soddisfatte altre condizioni prelimi-

Capitolo I
14
nari che resero possibili le sintesi molecolari nell’evoluzione del1’Universo fisico. La prima condizione consisteva nell’esistenza di specie atomiche dotate d’attività chimica e questa fu realizzata dalla nucleosintesi stellare. Occorreva poi che gli elementi, estremamente diluiti, fossero concentrati per aumentarne la probabilità di reazione. Occorreva infine che i luoghi ove i reagenti si concentravano, fossero durevolmente e al riparo da temperature troppo elevate e da radiazioni troppo intense. Solo in tal modo i prodotti di reazione avevano la pos-sibilità di accumularsi ed interagire ulteriormente fra loro. Gli ambien-ti in cui tutte queste condizioni si sono contemporaneamente verificate furono le zone periferiche delle nebulose in via di sedimentazione in-torno ad un astro centrale e, più particolarmente, i pianeti derivanti dalla loro condensazione.
Durante il processo d’accrescimento da frammenti e gas e, succes-sivamente, nella fase d’intenso bombardamento meteoritico delle su-perfici planetarie, vanno perduti la maggior parte dei componenti gas-sosi, ed in particolare quasi tutto l’elio e l’idrogeno non combinato. Questo degassamento è particolarmente spinto nelle zone relativamen-te più prossime all’astro centrale a causa della pressione di radiazione e della temperatura elevata. Alla fine del processo i pianeti interni ri-sultano dei concentrati di composti che si trovano quasi esclusivamen-te allo stato di fasi condensate.
In Tabella 1.2 riportiamo schematicamente la distribuzione dei principali elementi nel nostro pianeta ed alla sua superficie in fase so-lida e liquida. Affinché, possano essere sintetizzate molecole più complesse dei semplici composti inorganici, occorre che siano soddi-sfatte due ulteriori condizioni. Una consiste nella presenza di un am-biente gassoso stabile, o meglio ancora di un sistema termodinamico costituito da una fase liquida ed una gassosa in equilibrio. La seconda nell’esistenza in quest’ambiente di una consistente, ma non eccessiva, quantità di azioni ionizzanti. Entrambe queste condizioni si sarebbero verificate alla superficie del nostro pianeta qualche centinaio di milio-ni di anni dopo il completamento della fase di accrezione.

Fisica e Propedeutica Biofisica
15
Tabella 1.3. Permanenza medie di alcuni gas nell’atmosfera terrestre, in secondi. La temperatura media equivalente negli strati atmosferici superiori, da cui avviene la fuga, è nell’ordine dei 1000 K. (I dati sono tratti da Spitzer L Jr (1952) “The terre-strial atmosphere above 300 km”, pp. 211-247, in Kuiper GP (curatore), “The At-
mospheres of the Earth and Planets”, University of Chicago Press, Chicago).
Specie atomica considerata Temperatura
(K) Idrogeno Elio Azoto Ossigeno
300 1,7 × 1010 >1035 500 2,4 × 107 4,2 × 1025 1092 10105
1000 3,6 × 104 4,0 × 1013 1045 1051 2000 1,8 × 103 2,4 × 107 3,9 × 1022 1,0 × 1026
La produzione dell’atmosfera secondaria e dell’idrosfera furono dovute all’attività geologica. I principali prodotti gassosi emessi dai vulcani sono CO2, N2, SO2, H2, Cl2, SO3, ed il neon. La gravità terre-stre è risultata sufficientemente forte da trattenere la maggior parte di questi gas, come si vede in Tabella 1.3, dove vengono riportati i tempi di permanenza di alcuni gas nella nostra atmosfera. Un’altro fonda-mentale contributo del vulcanesimo consisteva nell’emissione di H2O, sia sotto forma di vapore che come acqua iuvenile emessa dal magma, che giungeva in superficie allo stato liquido. Oltre a formare a lungo andare un’idrosfera permanente, quest’acqua interagiva con le rocce superficiali, all’epoca quasi tutte costituite da minerali allo stato ridot-to, date le condizioni di formazione in presenza di eccesso di idroge-no. L’acqua agendo sui nitruri diede luogo a NH3 e N2, con i solfuri a H2S, con i carburi a CH4 ed ossido di carbonio. Al ritmo attuale dell’attività vulcanica si può valutare in alcune centinaia di milioni di anni il tempo richiesto per la formazione di un’atmosfera e di un idro-sfera stabili. Il dinamismo di quest’ambiente doveva comportare una rilevante attività elettrogenica, con produzione di scariche calde ed effetto corona, ambedue efficaci cause ionizzanti.

Capitolo I
16
Tabella 1.4. Sorgenti d’energia e loro contributi energetici alla superficie del suolo all’epoca dell’evoluzione chimica, in assenza di uno strato d’ozono, che blocca la
radiazione UV. La radiazione solare a lunghezze d’onda superiori a 200 nm è poco o per nulla efficace ai fini del meccanismo che produce le sintesi di tipo Miller.
Sorgente di energia Apporto di energia al suolo (kjoule/m2 anno)
Radiazione ultravioletta < 150 mn 150–200 nm 200–250 nm 250–300 nm
71 1.650
22.000 119.000
Radiazione solare totale 10.900.000 Scariche elettriche Effetto corona
Fulmini (scarica) Fulmini (onda d’urto)
126 42 46
Radioattività naturale da profondità 0 a 500 m estrapolato a 4,5 × 109 anni fa
60
Raggi cosmici e vento solare 8,1
Il decadimento dei radionuclidi e la radiazione solare — soprattutto la componente UV — costituivano altri fattori ionizzanti sinergici (Tabella 1.4). L’accumulo nel mare e nelle acque superficiali di pro-dotto molecolari derivanti dall’azione delle radiazioni ionizzanti sull’atmosfera primitiva ha portato alla formazione di quella “zuppa diluita” in cui le successive fasi evolutive, hanno trovati i “mattoni” con cui edificare strutture complesse ancora maggiori. Quindi, alcune centinaia di milioni di anni dopo la formazione dei pianeti, sulla Terra (e forse su Marte), erano presenti tutte le condizioni necessarie — ma non certamente sufficienti — alla sintesi di molecole organiche com-plesse. Una volta formate, queste molecole venivano dilavate nell’idrosfera che offriva riparo efficace da temperature troppo elevate e da dosi eccessive di radiazioni ionizzanti o da scariche elettriche.
Nel 1952, Harold C. Urey e Stanley L. Miller condussero una serie di esperimenti sugli effetti delle scariche elettriche in un’atmosfera riducente, simile a quella primitiva sulla Terra. Sottoponendo una mi-scela di idrogeno, metano, ammoniaca ed acqua a scariche elettriche nell’apparecchio rappresentato in Figura 1.6, essi osservarono proces-si di sintesi spontanea di aldeidi, acidi carbossilici ed aminoacidi.

Fisica e Propedeutica Biofisica
17
Figura 1.6. Apparecchiatura usata nell’esperimento originale di Urey–Miller.
In un tipico esperimento alla Miller il gas veniva fatto circolare nella zona sottoposta alla scarica elettrica per una settimana; il conte-nuto iniziale di carbonio, sotto forma di CH4 era di circa 0,06 mol e circa il 15% di questo carbonio veniva convertito in composti organi-ci, tra cui acido cianidrico, gli aminoacidi glicina, alanina, acido glu-tammico ed acido aspartico nonché idrocarburi ed un’altra ventina di molecole tra cui l’acido aspartico e quello α–amminoisobutirrico. Ap-portando alcune varianti sperimentali sono stati sintetizzati anche nu-merosissimi altri composti, fra i quali la maggior parte degli aminoa-cidi naturali e delle comuni basi azotate. Questi risultati hanno giu-stamente risvegliato un enorme interesse e costituiscono un indubbio progresso nella conoscenza dei meccanismi che hanno portato alla formazione spontanea di molecole complesse. L’apparecchiatura è riempita d’idrogeno, metano, ammoniaca ed acqua. La scarica elettri-

Capitolo I
18
ca fra elettrodi di tungsteno produce ioni che ricombinandosi fra loro formano molecole organiche, anche relativamente complesse. Questi prodotti vanno a concentrarsi nell’acqua contenuta nel pallone piccolo a sinistra in basso nella figura.
1.1.2.2. L’evoluzione prebiologica
Gli organismi viventi contengono macromolecole molto complesse, sia di tipo informazionale, come gli acidi nucleici, sia di tipo funzionale, come gli enzimi, costituite da migliaia o milioni di atomi collegati in architetture sofisticate e precise. Strutture d’ordine gerarchico superiore, come membrane, organelli, cellule ed organismi, coordinano l’azione di queste macromolecole e di un grandissimo numero di altre nell’espletamento di funzioni specifiche. Come si è potuto produrre un così alto grado di ordine strutturale e funzionale, quando l’evoluzione chimica riesce solo a generare la zuppa diluita entro cui sono mescolati alla rinfusa composti relativamente semplici e già la dimerizzazione incontra ostacoli tanto rilevanti?
Per comprendere come ciò sia potuto accadere occorre tener pre-sente un aspetto fin qui trascurato da quanti si sono occupati del pro-blema. Esso consiste nei sinergismi che si stabiliscono fra forze e strutture e nel progressivo cambiamento dei tipi di forze responsabili dell’evoluzione lungo i vari stadi del processo.
Già nelle fasi precedenti abbiamo osservato gli effetti di questo al-ternarsi di ruoli fra forze e strutture. La formazione delle grandi archi-tetture cosmiche aventi volumi caratteristici di 1070 metri cubi, — i superammassi galattici, le galassie e le stelle — è determinata unica-mente dall’interazione gravitazionale e dalle forze inerziali. Adden-sando la materia, esse creano le condizioni in cui l’interazione forte — operante su volumi inferiori ai primi di oltre cento ordini di grandezza — provoca la seconda nucleosintesi, e laddove la temperatura scende a sua volta di molti ordini di grandezza, l’interazione elettromagnetica può poi generare atomi e molecole.
Protagoniste della fase successiva, di evoluzione prebiologica, sono delle forze ancora diverse, il cui modus operandi dipende in modo sempre più evidente dal grado di organizzazione delle strutture su cui agiscono. Per meglio chiarire questo fondamentale concetto sarà op-

Fisica e Propedeutica Biofisica
19
portuno considerare brevemente il comportamento dei biopolimeri in ambiente acquoso dove i singoli aminoacidi di una proteina sono sog-getti come qualsiasi altra molecola alle normali interazioni con il mezzo circostante. La specificità della loro sequenza peraltro determi-na dei punti in cui la catena può collegarsi con se stessa, guidata dalle interazioni idrofi1ico–idrofobiche con il solvente. Ne risulta una spe-cifica conformazione tridimensionale: a fune nel collagene, a cane-strello abbracciante il sito attivo nella mioglobina, ecc. A questa strut-tura terziaria è legata la specificità funzionale del polimero. Le forze sono le stesse che operano su e fra ogni altro tipo di molecole in solu-zione, ad esempio sugli aminoacidi componenti; entro una tale solu-zione quelle forze non produrranno mai un’architettura ordinata. Inve-ce non appena la struttura macromolecolare con l’adatta successione di aminoacidi viene posta in una soluzione a pH e temperatura compa-tibili, la struttura tridimensionale si forma spontaneamente in una fra-zione di secondo.
Così più in generale, forze che producono effetti scarsi o nulli ad un determinato livello di organizzazione di un sistema, divengono de-terminanti quando altre forze hanno ordinato preventivamente il si-stema stesso.
Torniamo al problema prebiologico ed alla zuppa diluita prodotta nell’idrosfera terrestre dall’evoluzione chimica. Molti dei composti organici in essa contenuti sono scarsamente solubili ed il loro accumu-lo ne provoca la segregazione sotto forma di fase gelatinosa: goccioli-ne nel mare o straterelli depositati sulle rocce e sui fanghi del fondo.
La formazione di questo subsistema di aree gelatinose pre–vitali, chiamate da Pirie “eobionti” suddivide (Figura 1.7) l’ambiente prebio-logico in due compartimenti: gli eobionti e la soluzione acquosa circo-stante. Le molecole più idrofobiche sintetizzate nei processi alla Mil-ler si vanno a disporre spontaneamente all’interfaccia generando una rudimentale membrana. È facile convincersi che questi processi in cui viene creato dell’ordine strutturale macroscopico, sono in realtà gui-dati dall’entropia (entropy–driven) in quanto decorrono con aumento dell’entropia del sistema complessivo. A contatto con una superficie idrofobica l’acqua acquista un più alto grado d’ordine interno; la se-

Capitolo I
20
gregazione dei materiali idrofobici in una fase condensata, riducendo la superficie di contatto con l’acqua n’aumenta l’entropia.
Figura 1.7. Strati di gelatina depositati su rocce o fanghi e goccioline del materiale gelatinoso sospesi nel mare possono essere stati sottoposti a gradienti di temperatura per periodi prolungati. Le due situazioni schematicamente rappresentate si riferisco-no a strati alla Bernal attraversati da flussi di calore d’origine geotermica ed a coa-cervati d’Oparin inglobanti un granulo minerale che cattura la luce del Sole. Ad esempio supponendo: (rcoacervati/rgranuli) = 5 il flusso di calore in uscita risulterebbe venticinque volte minore di quello di energia radiante in arrivo dal Sole, Jo ed, es-sendo K la conducibilità termica della gelatina, il gradiente radiale di temperatura, dato da: )()/( 22
gco
gc rrKJrrT =−∆ risulta assai rilevante, quanto rcoacervati ≤ 1
micron.
La compartimentalizzazione della gelatina primitiva e la formazio-ne di una rudimentale membrana idrofobica sono processi spontanei, che hanno molteplici conseguenze: i composti idro– e liposolubili pas-seranno facilmente lo strato limite, mentre quelli più idrofili tenderan-no ad essere confinati nel mezzo acquoso interno od in quello esterno. Le molecole dotate di una parte idrofilica ed un’idrofobica rimarranno fissate allo strato limite, sporgendone fuori con l’estremo idrofilico. La membrana tenderà così col tempo ad acquistare una maggiore com-plessità.