
Aggiornamenti nella gestione dell’allevamento...
14
Percorso di formazione a distanza (FAD) abbinato a SUMMA ANIMALI DA REDDITO Aggiornamenti nella gestione dell’allevamento bovino Crediti ECM complessivi: 28 Validità: 15 marzo 2014 - 14 marzo 2015 www.pviformazione.it PRIMA PARTE Il complesso metrite-endometrite Autori: Giovanni Gnemmi, Elisa Ferrari, Cristina Maraboli Medici veterinari liberi professionisti, Bovinevet Bovine Ultrasound Services, Premosello Chiovenda (VB) SECONDA PARTE L’ingestione della bovina Autore: Alessandro Fantini Medico veterinario libero professionista, Dairy Production Medicine Specialist Anguillara Sabazia (RM) TERZA PARTE Le patologie dell’ovaio Autori: Giovanni Gnemmi, Elisa Ferrari, Cristina Maraboli Medici veterinari liberi professionisti, Bovinevet Bovine Ultrasound Services, Premosello Chiovenda (VB) QUARTA PARTE La nutrizione proteica e il bilanciamento amminoacidico Autore: Alessandro Fantini Medico veterinario libero professionista, Dairy Production Medicine Specialist Anguillara Sabazia (RM) QUINTA PARTE Il ruolo della formazione nella gestione riproduttiva Autori: Giovanni Gnemmi, Juan Manuel Ramos Medici veterinari liberi professionisti, Bovinevet Bovine Ultrasound Services, Premosello Chiovenda (VB) SESTA PARTE Stato dell’arte sulla mastite bovina da Prototheca spp. Autori: Clara Locatelli, Valerio Bronzo, Bianca Castiglioni, Antonio Casula, Paola Cremonesi, Paolo Moroni, Claudia Pollera, Nicola Rota, Licia Scaccabarozzi Università degli studi di Milano, Dipartimento di Scienze Veterinarie per la Salute, la Produzione Animale e la Sicurezza Alimentare SETTIMA PARTE L’apporto energetico Autore: Alessandro Fantini Medico veterinario libero professionista, Dairy Production Medicine Specialist Anguillara Sabazia (RM) OTTAVA PARTE Come scegliere e formare un capo stalla Il ruolo della formazione nella gestione riproduttiva Autori: Giovanni Gnemmi, Juan Manuel Ramos Medici veterinari liberi professionisti, Bovinevet Bovine Ultrasound Services, Premosello Chiovenda (VB) NONA PARTE I carboidrati non strutturali e l’endotossicosi ruminale Autore: Alessandro Fantini Medico veterinario libero professionista, Dairy Production Medicine Specialist Anguillara Sabazia (RM) RESPONSABILE SCIENTIFICO Giovanni Sali Direttore Centro Studi “San Francesco”, Piacenza Sponsor:
Transcript of Aggiornamenti nella gestione dell’allevamento...

Percorso di formazione a distanza (FAD)abbinato a SUMMA ANIMALI DA REDDITO
Aggiornamenti nella gestionedell’allevamento bovino
Crediti ECM complessivi: 28Validità: 15 marzo 2014 - 14 marzo 2015
www.pviformazione.it
PRIMA PARTE
Il complesso metrite-endometriteAutori: Giovanni Gnemmi, Elisa Ferrari, Cristina MaraboliMedici veterinari liberi professionisti, Bovinevet Bovine Ultrasound Services, Premosello Chiovenda (VB)
SECONDA PARTE
L’ingestione della bovina Autore: Alessandro FantiniMedico veterinario libero professionista, Dairy Production Medicine SpecialistAnguillara Sabazia (RM)
TERZA PARTE
Le patologie dell’ovaioAutori: Giovanni Gnemmi, Elisa Ferrari, Cristina MaraboliMedici veterinari liberi professionisti, Bovinevet Bovine Ultrasound Services, Premosello Chiovenda (VB)
QUARTA PARTE
La nutrizione proteica e il bilanciamento amminoacidico Autore: Alessandro FantiniMedico veterinario libero professionista, Dairy Production Medicine SpecialistAnguillara Sabazia (RM)
QUINTA PARTE
Il ruolo della formazione nella gestione riproduttiva Autori: Giovanni Gnemmi, Juan Manuel Ramos Medici veterinari liberi professionisti, Bovinevet Bovine Ultrasound Services, Premosello Chiovenda (VB)
SESTA PARTE
Stato dell’arte sulla mastite bovina da Prototheca spp.Autori: Clara Locatelli, Valerio Bronzo, Bianca Castiglioni,Antonio Casula, Paola Cremonesi, Paolo Moroni, Claudia Pollera, Nicola Rota, Licia ScaccabarozziUniversità degli studi di Milano, Dipartimento di Scienze Veterinarie per la Salute, la Produzione Animale e la Sicurezza Alimentare
SETTIMA PARTE
L’apporto energetico Autore: Alessandro FantiniMedico veterinario libero professionista, Dairy Production Medicine SpecialistAnguillara Sabazia (RM)
OTTAVA PARTE
Come scegliere e formare un capo stallaIl ruolo della formazione nella gestione riproduttiva Autori: Giovanni Gnemmi, Juan Manuel RamosMedici veterinari liberi professionisti, Bovinevet Bovine Ultrasound Services, Premosello Chiovenda (VB)
NONA PARTE
I carboidrati non strutturali e l’endotossicosi ruminale Autore: Alessandro FantiniMedico veterinario libero professionista, Dairy Production Medicine SpecialistAnguillara Sabazia (RM)
RESPONSABILE SCIENTIFICO
Giovanni SaliDirettore Centro Studi“San Francesco”, Piacenza
Sponsor:
07-07-cop_GA_00-00_FAD Ghisleni 26/09/14 15.38 Pagina 7

8 N° 8 Ottobre 2014ANIMALI DA REDDITO
Percorso FAD
Modalità operative
Chi sono i destinatari del corso FAD?
Il corso, accreditato presso la Conferenza Nazionale perla Formazione Continua, è rivolto alla categoria dei me-dici veterinari.
È importante essere abbonati a SUMMA Animali dareddito per accedere al corso FAD?
Per gli abbonati a SUMMA Animali da reddito sono pre-viste condizioni riservate e particolarmente vantag-giose.
Come si svolge il corso?
Il corso è composto da 9 dossier (materiale formati-vo) pubblicati in successione su SUMMA Animali dareddito a partire da gennaio/febbraio 2014 (SUMMAn. 1) e fino a dicembre 2014 (SUMMA n. 10). Soltan-to il numero monotematico in uscita a maggio 2014(SUMMA n. 4) NON conterrà alcun dossier riferito alcorso FAD.
Come si ottengono i crediti ECM?
Per ottenere i crediti ECM è necessario seguire questisemplici passaggi:
Lettura dei DossierI dossier pubblicati in successione sui numeri di SUM-MA Animali da reddito durante l’anno 2014 rappresen-tano il materiale formativo e di studio. Si presentanocome articoli scientifici, contraddistinti sulla paginada uno specifico richiamo al corso FAD. Sono consultabili anche in formato digitale, accedendoalla versione on line del periodico su www.pointvet.it.
Registrazione/Login su www.pviformazione.itL’utente deve attivare un account all’indirizzohttp://fad.pviformazione.it/accedi. L’operazione è gra-tuita e senza obbligo di acquisto. Naturalmente chi aves-se già un account su questa piattaforma NON deve crear-ne uno nuovo, ma può utilizzare quello esistente.
Acquisto del questionarioGli abbonati a SUMMA Animali da reddito possono ac-
quistare dall’account personale il SOLO questionariodi valutazione dell’apprendimento al prezzo riservatodi € 36,00 (IVA inclusa).
Attestato ECMSuperato il questionario di valutazione dell’apprendi-mento e compilato il questionario di valutazione dellaqualità percepita, è possibile dal proprio account ef-fettuare il download dell’attestato con i crediti ECM.A seconda della data di superamento del questionario,i crediti saranno attribuiti all’anno 2014 (corso con-cluso entro il 31.12.14) o all’anno 2015 (corso conclu-so dopo il 31.12.14).
Come è composto il questionario?
Il questionario verte sui temi trattati dai singoli dossierpubblicati su SUMMA Animali da reddito ed è disponi-bile soltanto on line. Si compone di 9 test in successione, attivati in con-temporanea con l’uscita del dossier a cui si riferiscono.L’ultimo test pubblicato sarà pertanto quello riferito aldossier di SUMMA Animali da reddito n. 10, dicembre2014.
Ogni test presenta una serie di domande a risposta qua-drupla e scelta singola. Per superare il singolo test è ne-cessario rispondere correttamente almeno all’80% del-le domande.
Per informazioni dettagliate sul funzionamento dei test,si rimanda alle modalità operative FAD sul sito www.pvi-formazione.it.
Il questionario di valutazione dell’apprendimento siconsidera concluso una volta superati tutti e 9 i singo-li test. Per accedere al download dell’attestato ECM sa-rà sufficiente a questo punto compilare il form di va-lutazione della qualità percepita.
Quando termina il corso?
La validità del corso abbinato a SUMMA Animali dareddito termina in data 14 febbraio 2015. Dopo la sca-denza NON sarà più possibile ottenere i relativi cre-diti ECM.
08-20_FAD_00-00_FAD Ghisleni 26/09/14 15.37 Pagina 8

!!
9N° 8 Ottobre 2014 ANIMALI DA REDDITO
nergia è uno delle parole mag-giormente utilizzate da chi allevae assiste le vacche da latte. Viene“tirata in ballo” dal buiatra se lafertilità non è ritenuta soddisfa-
cente (carenza) oppure se le vacche gra-vide sono troppo grasse (eccesso). Inol-tre, apportare energia alle bovine per ilmantenimento, la crescita, la produzio-ne e la riproduzione è la maggiore vocedei costi della produzione del latte. Perquesto e per molti altri motivi, è un temada approfondire. Propedeuticamente èbene comprendere il peculiare meccani-smo attraverso il quale la vacca da latteproduce l’energia chimica necessaria aisuoi, spesso esasperati, fabbisogni bio-logici. Una carenza energetica ha un sen-sibile impatto sulla produzione di latte,sulla fertilità, sulla piena efficienza delsistema immunitario e, comunque, sullalongevità funzionale della bovina. Peri-colosi sono anche gli eccessi energeticinei periodi in cui il rischio di bilancioenergetico negativo è inesistente, comedurante la gravidanza e fino alle ultimedue settimane prima del parto. In questoperiodo e soprattutto nelle bovine di al-to potenziale genetico, aumentano le pro-babilità di ingrassamento con tutto ciòche comporta come fattore di rischio perla chetosi, la lipidosi epatica e tutte le al-tre patologie ad esse correlate.
La produzione di energia1. Il calcolo dell’energiaLe equazioni di stima della concentra-zione energetica di un alimento o di unadieta per bovine da latte sono basate sulconcetto di energia netta, espressa comeEnergia Netta Lattazione (ENL) o UnitàForaggera Latte (UFL), ossia l’energia del-la razione disponibile per la produzionedi latte. Arrivare ad esprimere la con-centrazione di energia netta di un ali-mento o di una razione alimentare com-porta una serie di equazioni che partonodall’energia lorda, ossia dell’energia ter-mica sprigionata da un alimento “bru-
ciato” in una bomba calorimetrica, chemisura la quantità totale di energia chi-mica disponibile partendo da quella ter-mica. Dall’energia lorda si passa all’e-nergia digeribile, ossia a quella che ri-mane dopo la dispersione con le feci. L’in-gestione è il fattore animale che più in-fluenza la digeribilità dell’energia. L’ef-ficienza marginale di digeribilità dimi-nuisce come l’ingestione aumenta. L’e-nergia metabolizzabile è quella che ri-mane dopo le perdite di calorie con le uri-ne e il gas.Le diete fibrose aumentano la produzio-ne di metano e l’ingestione di molte pro-teine aumenta la sintesi dell’urea, pro-cesso che richiede un considerevole con-sumo energetico. Entrambi riducono l’ef-ficienza nella conversione dall’energia di-geribile a metabolizzabile.Le diete ricche di amido riducono la pro-duzione di metano e facilitano la con-versione.Infine, l’energia netta latte è l’energia se-creta nel latte o ritenuta nel corpo e usa-ta come mantenimento. L’energia netta èl’energia di mantenimento meno il calo-re disperso per l’inefficienza della tra-sformazione. Quando aumentano la fibrae le proteine della razione diminuisce l’ef-ficienza della trasformazione, mentrequando aumentano amidi e grassi, l’effi-cienza aumenta. A titolo di esempio, sipuò valutare il flusso energetico di unadieta destinata a una bovina di 600 kg cheproduce poco più di 30 kg di latte al gior-no. Delle 90 Mcal giornaliere ingerite, 27vengono disperse con le feci nel passag-gio a energia digeribile. Dalla digeribilealla netta ne vengono disperse 4 con leurine, 6 come gas e ben 21 come calore.Il risultato è che diventa energia netta so-lo il 35% di quella ingerita che, comun-que, è ben più elevata rispetto alla tra-sformazione di energia chimica a ener-gia meccanica di un motore a scoppio,dove il rendimento è solo del 15% (figu-ra 1).La stima dell’approvvigionamento ener-getico di una bovina da latte ha enormi
EL’apporto energeticoALIMENTAZIONE
Alessandro FantiniDairy Production Medicine Specialist, Fantini Professionale Advice srl, Anguillara Sabazia (Roma)
■ RIASSUNTO
La vacca da latte, ma come del restotutti i ruminanti, ha un suo peculiaremetodo per produrre l’energia chimicafondamentale per il suo mantenimento,la crescita, la riproduzione, le difeseimmunitarie e la produzione del latte.Solo la profonda conoscenza dei meccani-smi biochimici da essa utilizzati consenteal buiatra di trovare le soluzioni piùidonee alla gestione del bilancio energe-tico e proteico negativo. È anche difondamentale importanza la diagno-stica soggettiva e oggettiva, ossial’adozione di tutte le valutazioni perquantificare l’entità di questa condizionemolto pericolosa per la produzione, lasalute e la longevità produttiva dellavacca da latte. Molto pericolose sonole scelte alimentari o l’adozione diadditivi mutuate dai monogastrici perapportare energia alle bovine.
Parole chiave:Bilancio energetico,bilancio proteico, gluconeogenesi,vacca da latte.
■ SUMMARY
Energy intake
The dairy cow, like all ruminants, hasa peculiar method to produce chemicalenergy essential for growth, reproduc-tion, immunity and milk production.Only a deep knowledge of the underly-ing biochemical mechanisms allowsvets to find the most appropriatesolutions to manage negative proteinand energy balance. Subjective andobjective diagnostics is also crucial,namely the adoption of all assessmentsto quantify the extent of this verydangerous condition for production,health and productive longevity ofdairy cows. Very dangerous are nutritionalchoices or the use of additives frommonogastric to provide energy to cows.
Key words:energy balance, proteinbalance, gluconeogenesis, dairy cow.
Aggiornamenti nella gestione dell’allevamento bovino - DOSSIER N. 7
08-20_FAD_00-00_FAD Ghisleni 26/09/14 15.37 Pagina 9

limiti concettuali. Il sistema di calcolodell’energia serve solo ed esclusivamen-te a fornire un ordine di grandezza delproblema. Alla bovina, in realtà, serve l’e-nergia chimica che si sprigiona dalla con-versione della molecola ATP in ADP. Lapossibilità di correlare questo processochimico con il calcolo dell’energia trovadifficoltà soprattutto per le grandi pro-duttrici di latte. Un altro concetto fon-damentale è che la bovina è un ruminan-te, ossia trasforma buona parte degli ali-menti ingeriti in microrganismi rumina-li ed acidi grassi volatili. La quota di nu-trienti che supera il rumine non degra-data è comunque minoritaria, anche seimportante. Quello che interessa, per-tanto, è garantire alla bovina la massimaquantità possibile di ATP, soprattuttoquando il bilancio energetico è negativo.
2. Il ciclo di KrebsÈ bene ricordare che il focus deve esseresulla produzione di energia di un orga-nismo vivente che, nel nostro caso, è lavacca da latte. Diversa è la possibilità distima del valore energetico di un alimentoo di una razione alimentare. Sono ovvia-mente due aspetti altamente correlati manon sinonimi. Emblematica è la frase pro-nunciata da William P. Weiss, uno dei pa-dri dell’equazione più importante di cal-colo dell’energia e dei suoi fabbisogni,
che nel 2010 disse: “Il nutrizionista nondeve esitare ad aggiustare il modello chegenera il valore di ENL in base all’espe-rienza, alla composizione totale della die-ta e all’ingestione della bovina”.L’energia, che è semplicemente una mo-lecola presente all’interno dei mitocon-dri delle cellule, viene generata o meglio“caricata” in quello che viene chiamato ilciclo di Krebs, in nome di chi lo ha de-scritto nel 1937 e che gli valse nel 1953 ilpremio Nobel per la medicina. La pro-fonda conoscenza di questo ciclo, notoanche come il ciclo degli acidi tricarbos-silici e soprattutto delle molecole neces-sarie alla produzione dell’energia, è pro-pedeutica alla ricerca delle soluzioni perla gestione del bilancio energetico dellabovina e orienta il nutrizionista verso que-gli alimenti in grado di fornire il “com-bustibile” più appropriato per questa im-portante reazione biochimica. La cono-scenza del ciclo di Krebs si deve neces-sariamente integrare con la conoscenzadella biochimica ruminale: quali sono lemolecole funzionali alla produzione del-l’energia che può produrre il rumine e co-me avviene l’assorbimento intestinale deinutrienti?Il ciclo di Krebs è un ciclo metabolico fon-damentale in tutte le cellule che utiliz-zano ossigeno, ossia nel processo dellarespirazione cellulare. Negli organismi
Percorso FAD
10 N° 8 Ottobre 2014ANIMALI DA REDDITO
Aggiornamenti nella gestione dell’allevamento bovino
Figura 1. Flusso energetico della bovina in lattazionePeso vivo 600 kg – Produzione 31,7 kg
08-20_FAD_00-00_FAD Ghisleni 26/09/14 15.37 Pagina 10

!!
aerobici questo processo biochimico è l’a-nello di congiunzione delle vie metaboli-che responsabili della degradazione (ca-tabolismo) degli amminoacidi, dei car-boidrati e degli acidi grassi per la for-mazione dell’energia chimica. Il ciclo diKrebs fornisce anche molti precursori perla produzione di alcuni amminoacidi e dialtre molecole importanti per la cellula.Il luogo nel quale esso avviene è il mito-condrio. Il catabolismo del glucosio (gli-colisi) e lipidico (beta-ossidazione) pro-duce acetil-CoA. All’inizio si condensacon l’ossalacetato (forma ionizzata del-l’acido ossalacetico) per produrre il ci-trato. L’ ossalacetato è anche il punto diarrivo e di partenza per la demolizione ela sintesi degli amminoacidi, come l’aci-do aspartico e l’asparagina. Inoltre, l’a-spartato sintetizzato a partire dall’ossa-lacetato interviene nel ciclo dell’urea edel citrato, in quanto serve a immetterein questo ciclo uno ione ammonio, le-gandosi alla citrullina e formando argi-nin-succinato. Conosciamo l’estrema im-portanza che, soprattutto nei ruminanti,ha il ciclo dell’urea. Al termine del ciclodi Krebs, i due atomi di carbonio immessinell’acetil-CoA vengono ossidati in duemolecole di CO2, rigenerando completa-mente l’ossalacetato. La parte più im-portante di questo processo è la produ-
zione della molecola di GTP, che vieneimmediatamente utilizzata per rigenera-re una molecola di ATP. I cofattori NADHe FADH2, quando ridotti, sono in gradodi trasportare energia fino alla catena re-spiratoria mitocondriale. Presso tale ca-tena vengono riossidati in NAD+ e FAD ecedono elettroni in grado di rigeneraremolecole di ATP dall’ADP. L’energia chesi ricava dalla completa demolizione del-la molecola di glucosio è di circa 28 mo-lecole di ATP (figura 2).Ai fini di una profonda comprensione delmeccanismo di produzione dell’energiadeve essere molto chiaro cosa si trova amonte del ciclo di Krebs. Questo è il se-condo stadio del catabolismo dei carboi-drati. La glicolisi degrada il glucosio inpiruvato, che viene trasferito dal cito-plasma nei mitocondri e convertito in ace-til-CoA. Le proteine vengono invece tra-sformate in amminoacidi. Alcuni di que-sti possono costituire una fonte energe-tica, poiché convertiti in alcuni interme-di del ciclo di Krebs, come la valina, l’i-soleucina e l’aspartato. Altri amminoaci-di, attraverso la gluconeogenesi, come adesempio avviene per l’alanina, servonoper produrre glucosio. Durante il bilan-cio energetico negativo, la bovina ricor-re alle sue riserve lipidiche mobilizzan-do i trigliceridi contenuti nel tessuto adi-
11N° 8 Ottobre 2014 ANIMALI DA REDDITO
DOSSIER N. 7 - Alimentazione
Figura 2. Ciclo degli acidi tricarbossilici o ciclo dell’acido citrico o ciclo di Krebs
08-20_FAD_00-00_FAD Ghisleni 26/09/14 15.37 Pagina 11

poso. Questi vengono idrolizzati dalle li-pasi in acidi grassi e glicerolo. Il glice-rolo entra nella glicolisi o può essere tra-sformato in glucosio nella via della glu-coneogenesi, mentre gli acidi grassi ven-gono degradati nella beta-ossidazione perpoi produrre acetil-CoA.
Le principali molecolenecessarie alla produzionedi energia nel ciclo di Krebs1. Catabolismo degli acidi grassiGli acidi grassi vengono stoccati princi-palmente nel tessuto adiposo come ri-serva energetica da utilizzarsi negli ulti-mi giorni di gravidanza e nelle prime set-timane di lattazione, quando il bilancioenergetico diventa negativo, ossia quan-do l’energia spesa essenzialmente per laproduzione di latte è superiore alla ca-pacità della bovina di ingerire la giustaquantità di nutrienti necessari. Si vedràsuccessivamente che una rilevante quan-tità di acidi grassi liberati dal tessuto adi-poso nel corso del NEBAL vengono cap-tati dalla mammella per la produzione delgrasso del latte e, quindi, dirottati dallaproduzione di energia nel ciclo di Krebs.Quando arriva al tessuto l’informazioneendocrina dell’insulina che segnala un’in-sufficiente disponibilità di glucosio ema-tico per la produzione di energia inizia lalipomobilizzazione. Le lipasi del tessutoadiposo idrolizzano i trigliceridi stocca-ti in acidi grassi e glicerolo. Il glicerolo,attraverso la formazione di glicerofosfa-to e di fosfogliceraldeide, segue le tappedella glicolisi per poi partecipare al ciclodi Krebs.Oltre agli acidi grassi provenienti dal tes-suto adiposo, sono disponibili per il ciclodi Krebs quelli assorbiti dall’intestino e,quindi, di provenienza alimentare e quel-li a corta catena di provenienza rumina-le, come l’acido acetico e l’acido butirri-co. Nel predisporre i piani alimentari perle bovine in NEBAL, il nutrizionista dà lapriorità alla produzione ruminale di aci-do propionico in quanto precursore pri-mario, nella gluconeogenesi, del glucosio,che rappresenta il metabolita più impor-tante. Regolando verso il basso il pH ru-minale, per favorire il massimo tasso dicrescita della flora microbica amilolitica,che è la principale responsabile della pro-duzione del propionato, si riduce di fattola produzione ruminale di acetato e bu-tirrato da parte dei batteri cellulosolitici.Pertanto, si ricorre agli acidi grassi con-
tenuti in alimenti come le oleaginose in-tegrali (semi di soia e di cotone) oppurea miscele di acidi grassi a lunga catena re-si inerti nel rumine attraverso processi in-dustriali di idrogenazione e saponifica-zione. Questi acidi grassi sono maggior-mente a lunga catena, ossia con un nu-mero di atomi di carbonio superiore a 16.Nell’intestino tenue arrivano acidi grassilegati al glicerolo sotto forma, in genere,di trigliceridi. Per la demolizione dei tri-gliceridi serve l’azione combinata dellabile con le lipasi pancratiche. La bile vie-ne prodotta dal fegato ed è un fluido com-plesso fatto di acqua, elettroliti, acidi bi-liari, colesterolo, fosfolipidi e bilirubina.Gli acidi biliari aiutano la digestione deigrassi. Le lipasi pancratiche rompono illegame degli acidi grassi con il glicerolorendendo possibile il loro assorbimento.Esistono dei limiti nell’uso degli acidi gras-si nella dieta della vacca da latte, legatinon tanto alle possibilità di assorbimen-to intestinale quanto nel tipo di fonte uti-lizzata. Gli oli liberi sono tossici per il ru-mine, mentre quelli insaturi potrebberosviluppare intermedi che, se assorbiti nel-l’intestino, possono inibire la sintesi digrasso del latte. Una dieta per vacche dalatte non dovrebbe contenere più del 2-2,5% di acidi grassi insaturi ossia non piùdi 500 g al giorno. Una parte dell’acido li-noleico (C18:2), contenuto principalmentenel seme di soia, nel mais e nel cotone, edi quello linolenico (C18:3), presente prin-cipalmente nel lino integrale, viene bio-idrogenato nel rumine con il rischio di svi-luppo di intermedi come il C18:1 trans 10,cis12 che, una volta assorbito a livello in-testinale, può inibire la sintesi del grassodel latte. È bene che non giungano più di70 g di questo acido grasso nell’intestino.In ogni caso, esiste un limite nell’impie-go degli acidi grassi nella dieta della vac-ca da latte. Tale limite è il 6% totale, os-sia, per una dieta che apporta 23, 5 kg disostanza secca, 1.400 g al giorno.Le tappe ossidative che servono a tra-sformare gli acidi grassi fino ad anidridecarbonica e acqua avvengono nei mito-condri. L’attivazione dell’acido grasso av-viene sulla membrana esterna del mito-condrio. Impegna Mg2+ e ATP ed è cata-lizzata dall’enzima acil-CoA sintetasi. L’a-cil-CoA così formato, per essere β-ossi-dato, deve essere trasportato nel mito-condrio. Qui di fondamentale importan-za è la carnitina, in quanto l’enzima car-nitina-aciltransferasi si esterifica con l’a-cido grasso per formare l’acil-carnitina.In questo modo l’acido grasso può entra-
Percorso FAD
12 N° 8 Ottobre 2014ANIMALI DA REDDITO
Aggiornamenti nella gestione dell’allevamento bovino
08-20_FAD_00-00_FAD Ghisleni 26/09/14 15.37 Pagina 12

!!
re nel mitocondrio per essere ossidato (fi-gura 3). Si evince pertanto che la carni-tina riveste un ruolo fondamentale nellacomplessa catena per la produzione del-l’energia. Una scarsa disponibilità di car-nitina non consente il passaggio nei mi-tocondri degli acidi grassi e quindi cau-sa un conseguente loro accumulo nel ci-toplasma delle cellule. Ben si sa che quan-do la bovina è in NEBAL, una rilevantequantità di acidi grassi arrivano alle cel-lule epatiche. Se nel citoplasma si accu-mulano gli acidi grassi, questi si posso-no riesterificare in trigliceridi e accu-mularsi nelle cellule epatiche dando luo-go, se la quantità è eccessiva, a lipidosiepatica. Questo accumulo patologico ditrigliceridi nelle cellule epatiche com-promette molte delle funzioni biochimi-che presiedute dal fegato, come la detos-sificazione dall’ammoniaca e dalle tossi-ne, la clerance degli ormoni steroidei, lostoccaggio di glicogeno ecc. Pertanto, lalipidosi epatica vede tra i suoi fattori ezio-logici l’eccessivo ingresso di acidi grassinella cellula epatica, ma anche una ca-renza relativa di carnitina.Vista l’importanza che ha la carnitina nelciclo di produzione dell’energia e nell’e-ziologia della lipidosi epatica deve esse-re meglio conosciuta, soprattutto perché
può essere utilizzata come additivo nel-l’alimentazione della vacca da latte. Lacarnitina è un acido carbossilico a cortacatena presente nei tessuti animali e inmodesta quantità nelle piante. Si chiamacosì perché fu estratta la prima volta nel1905 dalla carne bovina. Pur essendo strut-turalmente simile a un aminoacido, lacarnitina non forma proteine. La vaccada latte è in grado di sintetizzarla par-tendo da amminoacidi come la lisina e lametionina. Nell’uomo, è stata descrittauna malattia genetica che causa un’inca-pacità di sintetizzarla per una mutazio-ne genetica a carico delle proteina OCTN2.Nell’uomo, l’alimento che apporta pri-mariamente la carnitina è la carne bovi-na. Nella bovina, l’approvvigionamentoprimario è la sintesi a partire dalla me-tionina e dalla lisina. Si deve inoltre con-siderare che la L-carnitina è utilizzata nelpaziente diabetico in quanto migliora l’u-tilizzo di glucosio da parte dei tessuti,mediante l’azione indiretta di prevenzio-ne dell’accumulo di trigliceridi nelle cel-lule, che favorisce nettamente l’insulino-resistenza. È ben noto che l’insulino-ca-renza e l’insulino-resistenza rappresen-tano due condizioni parafisiologiche del-le bovine nelle ultime settimane di gra-vidanza e nelle prime settimane di latta-
13N° 8 Ottobre 2014 ANIMALI DA REDDITO
DOSSIER N. 7 - Alimentazione
Figura 3. β-ossidazione dei NEFA e chetogenesi nel fegato
08-20_FAD_00-00_FAD Ghisleni 26/09/14 15.37 Pagina 13

zione.La sintesi della carnitina avviene princi-palmente a carico della metionina, ami-noacido considerato essenziale nella vac-ca da latte. La metionina, che viene as-sorbita dall’intestino, deriva in parte dal-la biomassa batterica ruminale e in par-te dagli alimenti che ne sono ricchi, co-me il mais e suoi derivati, che riescono apassare indenni dal rumine e arrivare nel-l’intestino. Inoltre, può essere presentecome additivo anche in preparazioni ingrado di proteggerla dalla degradazioneruminale. Quindi, il pool di metioninadisponibile per la bovina può essere po-tenzialmente piuttosto elevato, quantifi-cabile nell ’ordine di diverse decine digrammi giornalieri. Nei primi 100 giornidi produzione la quantità di latte è mol-to elevata e con essa la produzione gior-naliera di caseina. Basti pensare che inquesto periodo una bovina può avere unaproduzione media di 40 kg di latte, conpunte di oltre 55 kg. Una bovina che pro-duce 40 kg di latte al 3,3% di proteina pro-duce ogni giorno 1.320 g di proteina, checorrisponde a circa un chilogrammo dicaseina. La caseina ha una concentra-zione di metionina pari al 2,5%, per cuicon il latte ne viene eliminata una quan-tità rilevante. Inoltre, la metionina è unaminoacido essenziale per la sintesi del-la caseina a livello mammario ma è an-che condizionante la caseificabilità, le-gandosi con la chimosina (caglio). Se l’a-limentazione destinata alle vacche fre-sche non assicura un apporto adeguatodi metionina con la proteina metaboliz-zabile e in considerazione della prioritàmetabolica di produrre latte, si possonoverificare carenze secondarie di questoaminoacido essenziale, difficilmente dia-gnosticabili con la sola riduzione dellaproteina percentuale del latte. Un caren-za di metionina è un fattore limitante lasintesi sia della carnitina sia della coli-na. Esiste un legame importante tra que-ste tre molecole, ossia metionina, carni-tina e colina, nell’eziologia della lipido-si epatica e, quindi, nell’utilizzazione ot-timale degli acidi grassi che arrivano inmammella. Più del 30% della colina vie-ne sintetizzata dalla metionina. Un ca-renza di colina limita la sintesi di carni-tina. In conclusione, si può affermare cheuna carenza di carnitina può fortementelimitare l’uso degli acidi grassi come “com-bustibile” del ciclo di Krebs in un mo-mento dove il bilancio energetico è ne-gativo e buona parte del glucosio che de-riva o dalla gluconeogenesi o dalle riser-
ve di glicogeno o dall’assorbimento inte-stinale viene prioritariamente reclutatodalla mammella per la sintesi della ca-seina e, quindi, del latte.
2. Il glucosioIl glucosio rappresenta il “combustibi-le” primario per la produzione di ATP e,quindi, di energia chimica delle cellule.È necessario ricordare che la vacca dalatte è un animale tendenzialmente ipo-glicemico avendo una glicemia normaleall’incirca di 40-60 mg/dl, pari a circa lametà di quella normale di un monoga-strico. Molto del pool di glucosio emati-co serve alla sintesi del lattosio e, quin-di, del latte. Per questo i tessuti dei ru-minanti sono molto tolleranti all’ipogli-cemia. La selezione genetica per produrresempre più latte e sempre più grasso hapremiato assetti ormonali riconducibilial diabete tipo 1 (insulino-carente) e ti-po 2 (insulino-resistente). Come è noto,la produzione di latte in una bovina hala priorità metabolica assoluta ed è fi-nalizzata ad aumentare al massimo la ca-pacità di sopravvivenza del vitello. Peravere la più elevata disponibilità di nu-trienti alla mammella sono stati “pre-miati” soggetti con una maggiore capa-cità di produrre GH, ossia l’ormone checrea quelle condizioni di un maggior af-flusso di nutrienti ai tessuti. Nel con-tempo, sono state premiate anche le bo-vine che hanno una minore reazione al-l’aumento della glicemia e quindi con unassetto endocrino riconducibile al dia-bete tipo 1. Essendo l’epitelio mamma-rio un tessuto pressoché privo di recet-tori per l’insulina, ne trae grande van-taggio nella captazione del glucosio pro-prio grazie a questo assetto ormonale. Adifferenza dell’epitelio mammario, il tes-suto adiposo ha molti recettori per l’in-sulina. L’ipoglicemia, indotta non tantodall’efficiente produzione di insulina pan-creatica ma dalla sottrazione di gluco-sio dalla mammella, stimola la libera-zione di acidi grassi dal tessuto adiposo,proprio per fornire al ciclo di Krebs unmetabolita alternativo al glucosio. È ti-pico delle vacche di alto potenziale ge-netico l’abilità di dimagrire vistosamen-te nelle prime settimane di lattazione.Questa liberazione, spesso imponente,di acidi grassi dà un ulteriore vantaggioselettivo, in quando queste molecole so-no anch’esse reclutate dalla mammellaper la sintesi del grasso del latte. Sem-pre con la finalità di dare alla mammel-la la massima quantità possibile di glu-
Percorso FAD
14 N° 8 Ottobre 2014ANIMALI DA REDDITO
Aggiornamenti nella gestione dell’allevamento bovino
08-20_FAD_00-00_FAD Ghisleni 26/09/14 15.37 Pagina 14
!!
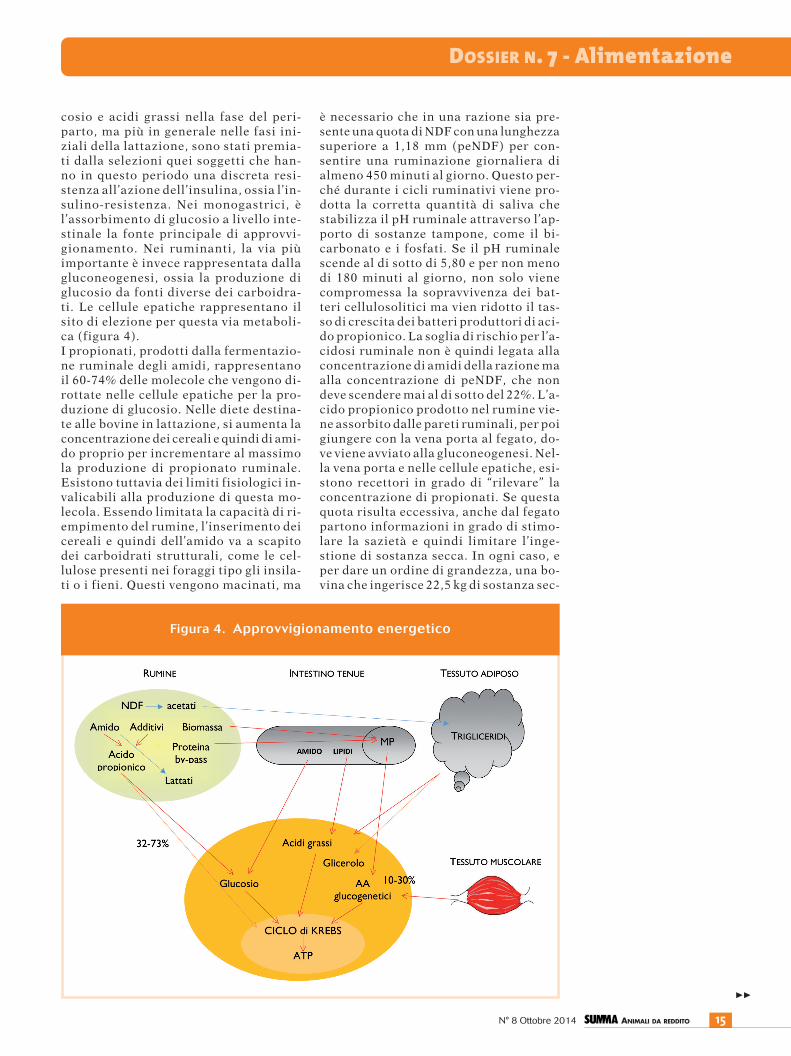
cosio e acidi grassi nella fase del peri-parto, ma più in generale nelle fasi ini-ziali della lattazione, sono stati premia-ti dalla selezioni quei soggetti che han-no in questo periodo una discreta resi-stenza all’azione dell’insulina, ossia l’in-sulino-resistenza. Nei monogastrici, èl’assorbimento di glucosio a livello inte-stinale la fonte principale di approvvi-gionamento. Nei ruminanti, la via piùimportante è invece rappresentata dallagluconeogenesi, ossia la produzione diglucosio da fonti diverse dei carboidra-ti. Le cellule epatiche rappresentano ilsito di elezione per questa via metaboli-ca (figura 4).I propionati, prodotti dalla fermentazio-ne ruminale degli amidi, rappresentanoil 60-74% delle molecole che vengono di-rottate nelle cellule epatiche per la pro-duzione di glucosio. Nelle diete destina-te alle bovine in lattazione, si aumenta laconcentrazione dei cereali e quindi di ami-do proprio per incrementare al massimola produzione di propionato ruminale.Esistono tuttavia dei limiti fisiologici in-valicabili alla produzione di questa mo-lecola. Essendo limitata la capacità di ri-empimento del rumine, l’inserimento deicereali e quindi dell’amido va a scapitodei carboidrati strutturali, come le cel-lulose presenti nei foraggi tipo gli insila-ti o i fieni. Questi vengono macinati, ma
è necessario che in una razione sia pre-sente una quota di NDF con una lunghezzasuperiore a 1,18 mm (peNDF) per con-sentire una ruminazione giornaliera dialmeno 450 minuti al giorno. Questo per-ché durante i cicli ruminativi viene pro-dotta la corretta quantità di saliva chestabilizza il pH ruminale attraverso l’ap-porto di sostanze tampone, come il bi-carbonato e i fosfati. Se il pH ruminalescende al di sotto di 5,80 e per non menodi 180 minuti al giorno, non solo vienecompromessa la sopravvivenza dei bat-teri cellulosolitici ma vien ridotto il tas-so di crescita dei batteri produttori di aci-do propionico. La soglia di rischio per l’a-cidosi ruminale non è quindi legata allaconcentrazione di amidi della razione maalla concentrazione di peNDF, che nondeve scendere mai al di sotto del 22%. L’a-cido propionico prodotto nel rumine vie-ne assorbito dalle pareti ruminali, per poigiungere con la vena porta al fegato, do-ve viene avviato alla gluconeogenesi. Nel-la vena porta e nelle cellule epatiche, esi-stono recettori in grado di “rilevare” laconcentrazione di propionati. Se questaquota risulta eccessiva, anche dal fegatopartono informazioni in grado di stimo-lare la sazietà e quindi limitare l’inge-stione di sostanza secca. In ogni caso, eper dare un ordine di grandezza, una bo-vina che ingerisce 22,5 kg di sostanza sec-
15N° 8 Ottobre 2014 ANIMALI DA REDDITO
DOSSIER N. 7 - Alimentazione
Figura 4. Approvvigionamento energetico
08-20_FAD_00-00_FAD Ghisleni 26/09/14 15.37 Pagina 15

ca, può produrre anche 7.600 g di acidigrassi volatili e di questa quota l’acidopropionico rappresenta una percentualeconsiderevole.Altra molecola importante nella gluco-neogenesi è il lattato. Questo può rap-presentare il 16-26% dei precursori delglucosio nella gluconeogenesi. Il lattatopuò derivare dalla fermentazione rumi-nale dei carboidrati non strutturali. Que-sta molecola deriva maggiormente dalcatabolismo del glucosio a carico del tes-suto muscolare, dell’utero e della pla-centa, nella fase più avanzata della gra-vidanza. Alcuni amminoacidi glucoge-netici hanno nella gluconeogenesi un ruo-lo fondamentale, in quanto contribui-scono dall’8 all’11% alla sintesi ex novodel glucosio, con punte fino al 26% al pic-co di lattazione. 100 grammi di ammi-noacidi glucogenetici possono produrre58 g di glucosio. Hanno potenzialità gluconeogenetica siagli amminoacidi essenziali, come la me-tionina, il triptofano, la treonina e la va-lina sia, e soprattutto, quelli non essen-ziali, come la glicina, l’acido glutammi-co, la prolina, la serina, l’acido asparti-co, l’idrossiprolina, la cistina, l’istidinae l ’alanina. Quest ’ultimo può rappre-sentare il 3-5% del totale dei precursoriper la gluconeogenesi. Durante il ciclodi Krebs, possono essere sintetizzati exnovo amminoacidi a partire dal piruva-to, l’ossalacetato e l’α-chetoglutarato. Ilglicerolo, che viene maggiormente libe-rato nella lipolisi, ossia dai trigliceridi,viene utilizzato nella bovina come pre-cursore del glucosio arrivando a rap-presentare per la gluconeogenesi lo 0,5-6% del totale (figura 5 e tabella 1).
Oltre al glucosio prodotto nella gluco-neogenesi esiste quello che può essereassorbito dalla bovina a livello intesti-nale dagli amidi che riescono a raggiun-gere l’intestino tenue perché dotati dibassa degradabilità ruminale. Questaquota dipende sia dalle caratteristichechimico-fisiche del cereale di provenienzasia dalla velocità di transito della razio-ne. A parità di inclusione nella razione,se la razione ha una quota totale di con-centrati piuttosto elevata la quantità diamido che può arrivare nell’intestino èpiù elevata. Di converso, esistono cerealicome il mais che, grazie alla conforma-zione dei granuli dell’amido, ha una de-gradabilità ruminale più bassa. Più ingenerale, nell’intestino tenue possono ar-rivare sia amidi sia saccarosio. Il pan-creas produce un enzima, l’a-amilasi, chedemolisce gli amidi in maltosio, malto-triosio e destrine, che sono però mole-cole troppo complesse per essere assor-bite dagli enterociti. Queste cellule in-testinali producono enzimi come malta-si, lattasi e saccarasi, che trasformanole molecole più complesse in glucosio,saccarosio e fruttosio così da essere as-sorbite.Esistono dei limiti fisiologici alla possi-bilità della vacca da latte di “processare”i carboidrati non strutturali nell’intesti-no tenue. Tali limiti non sono completa-mente conosciuti. L’abuso di amido bypass può causare un massiccio ingressonel colon e ciò può causare l’acidosi in-testinale. Si deve comunque rilevare cheanche il grosso intestino ha un ruolo noncosì marginale nell’approvvigionamentoenergetico della vacca da latte. Il 95% de-gli acidi grassi volatili prodotti nel colon
16 N° 8 Ottobre 2014ANIMALI DA REDDITO
Figura 5. Produzione epatica di glucosio
Percorso FAD Aggiornamenti nella gestione dell’allevamento bovino
08-20_FAD_00-00_FAD Ghisleni 26/09/14 15.37 Pagina 16

vengono assorbiti e ciò può rappresenta-re fino all’8-17% del totale degli AGV pro-dotti.
3. Gli amminoacidiEsistono degli amminoacidi che entranodirettamente nel ciclo di Krebs. Il più im-portante è l’alanina che, nel fegato, puòessere direttamente trasformata in piru-vato.Altri amminoacidi, come l’aspartato, lafenilalanina, la valina, l’isoleucina, la me-tionina e la tirosina sono coinvolti nellagluconeogenesi e nel ciclo di Krebs. Diconverso, da alcune molecole intermediedel ciclo di Krebs possono essere sinte-tizzati amminoacidi.Non marginale è il ruolo che hanno al-cuni amminoacidi nella gluconeogenesicome la glutammina, l’aspartato e la pro-lina. Gli amminoacidi glucogenetici so-no 13, di cui molti non essenziali in quan-to la bovina potrebbe sintetizzarli a par-tire da altri precursori. È difficile imma-ginare che la bovina non riesca a produrrepiù di 700 mmol/h di glucosio durante ilpicco produttivo (più di 3 kg al giorno)per i limiti derivanti dalla fisiologia delrumine. Di grande aiuto, oltre all’uso di un addi-tivo importante come il glicole propile-nico, è una maggiore cura dell’alimenta-zione proteica. Alcuni degli amminoaci-di glucogenetici, come la glutammina, l’a-
sparagina e la prolina, sono assorbiti da-gli organi drenati dalla vena porta e pos-sono dare un significativo contributo almantenimento del pool del glucosio. Neimomenti più “delicati” del bilancio ener-getico, ossia durante la fase di transizio-ne, l’imponente mobilizzazione di pro-teine labili accumulate in asciutta servea garantire l’apporto di amminoacidi ne-cessari per mantenere la produzione del-la proteina del latte, più che per la glice-mia. Di fondamentale importanza tra gli am-minoacidi non essenziali è l’alanina. Digrande interesse per i ricercatori, dap-
17N° 8 Ottobre 2014 ANIMALI DA REDDITO
DOSSIER N. 7 - Alimentazione
Giorni relativi al parto -19 -11 11 22 33 83Rilascio di glucosio mol/die 7 7,5 15,3 18,2 19,4 20,3Uptake lattato mol/die 2,2 3,1 6,4 5,7 5,5 3,4Contributo massimo (%) 15,7 20.7 20.9 15,6 14,2 8,4Uptake propionati mol/die 9,1 9,2 16,2 19,5 22,4 25,1Contributo massimo (%) 65 61,3 52,9 53,6 57,7 61,8Glicerolo mol/die 1 1 3 2,4 2 1,4Contributo massimo (%) 7,5 7 9,6 6,6 5,2 3,5AminoacidiContributo minimo (%) 11,8 11,3 16,5 23,8 22,9 26,3ProteineContributo minimo (g/die) 256 263 783 1.344 1.378 1.657
TABELLA 1. Contributo di diverse molecolealla gluconeogenesi in funzione del parto
Figura 6. Gli aminoacidi e il ciclo di Krebs
08-20_FAD_00-00_FAD Ghisleni 26/09/14 15.37 Pagina 17

prima, e per i tecnici, poi, è la possibili-tà di stimolare adeguatamente la gluco-neogenesi (figura 6).
Il bilancio energeticonegativoIl bilancio energetico e proteico negativodella vacca da latte negli ultimi giorni digravidanza è un evento fisiologico. Si èsempre indecisi se parlare del solo bilan-cio energetico negativo, in quanto duran-te questo lungo periodo si verificano an-che forti carenze amminoacidiche, moltedelle quali dovute al dirottamento degliamminoacidi verso la gluconeogenesi emaggiormente nella sintesi della caseina.Le ragioni sono diverse. Negli ultimi 60giorni di gravidanza, il feto acquisisce il65% del peso che avrà al momento del par-to. All’approssimarsi al parto viene rico-struito il parenchima mammario e iniziala produzione di colostro. Questi eventi fi-siologici sono però accompagnati da un ca-lo di ingestione, che ha anch’esso molte-plici ragioni e che può aggravarsi da fatticoncomitanti, come l’eccessivo ingrassa-mento delle bovine, stress sociali dovuti alrimescolamento degli animali ed errori neipiani alimentari o adozione di alimenti oadditivi scarsamente appetibili. Ad aggra-vare il tutto uno status di insulino-resi-stenza che investe periparto e puerperio,che ha come obiettivo fisiologico quello diaumentare ulteriormente la disponibilitàdi glucosio alla mammella per la sintesi dellattosio. Dopo il parto, il NEBAL diventafortemente negativo sempre a causa di unacapacità di ingestione dei nutrienti netta-
mente al disotto dei fabbisogni di glucosionecessari alla sintesi del lattosio mamma-rio e, quindi, del latte. La bovina, duranteil NEBAL, cerca di attenuare questo statusmobilizzando le riserve dapprima glucidi-che (glicogeno), poi amminoacidiche e in-fine lipidiche. La vacca da latte, una voltagravida, riesce con facilità a ripristinare lesue riserve lipidiche e spesso lo fa in mo-do eccessivo se l’alimentazione della se-conda metà della lattazione è eccessiva-mente energetica. Infatti, il problema del-le vacche che arrivano troppo grasse al mo-mento del parto è estremamente diffuso.Diversa è la possibilità che accumuli glu-cosio sotto forma di glicogeno e ammi-noacidi glucogenetici in quel complessodenominato “proteine labili”, ossia nonstrutturali, alle quali la bovina può ricor-rere per urgenti necessità energetiche. Do-po il parto, a causa del brusco abbassa-mento dell’insulina vengono rapidamentemobilizzate le riserve sia di glucosio sia diamminoacidi glucogenetici. Queste scor-te durano poco e comunque non oltre la fa-se del puerperio. Da ciò si evince che è ne-cessario somministrare alle bovine una ra-zione di asciutta molto equilibrata e nellafase finale della gravidanza evitare un pre-coce inizio di bilancio energetico e pro-teico negativo (tabella 2).La status energetico e proteico è fortemen-te “sorvegliato” dai sensori metabolici pre-senti sia a livello ipotalamico sia follicola-re e che hanno la funzione, tra le tante, diconsentire alla bovina di “prendere la deci-sione di riprodursi”, ossia ripristinare, allafine del puerperio, una corretta ciclicità deicicli estrali e avviare una nuova gravidanza
18 N° 8 Ottobre 2014ANIMALI DA REDDITO
% del fabbisogno energetico apportato 8 settimane dopo il parto ~ 95% McNamara (2002), Setter e Beever (2000)BCS al dry-off 2,75 Domecq (1997)BCS al parto 3,00 Mayne (2002), Hayirli (2002), Buckley (2003)% di vacche con perdita di più di 0,5 punti di BCS all’inizio della lattazione < 25% Buckley (2003), Pryce ( 2001) BCS alla fecondazione > 2,5 Pryce (2001), Buckley (2003)% di vacche a inizio lattazione con grasso/proteine > 1,5 < 10% Heuer (1999, 2000)% di vacche a inizio lattazione con nadir delle proteine < 3,05% < 15% Heuer (2000, Mayne (2002)% di vacche a inizio lattazione con un nadir di lattosio < 4,5% <15% Heuer (2000) Buckley (2003)Declino settimanale nella lattazione dopo il picco < 2,5% Chamberlain e Wilkinson(2002)Spazio in mangiatoia riservato alle vacche in transizione 0,6 m Grant e Albright ( 1995), Shaker (1993)Percentuale di scarto accettato nella mangiatoia della transizione > 3% Grant e Albright (1995), Robinson (1989)% di vacche 2-14 gg prima del parto con BHBA > 0,6 mmol/l < 10% Oetzel (2004), Whitaker (1997)% di vacche 2-14 gg prima del parto con NEFA > 0,4 mmol/l < 10% Oetzel (2004), Whitaker (1997)% di vacche a inizio lattazione con BHBA > 1,4 mmol/l < 10% Oetzel (2004), Whitaker (1997)% di vacche a inizio lattazione con NEFA > 0,7 mmol/l < 10% Oetzel (2004), Whitaker (1997)
TABELLA 2. Criteri di valutazione del NEBAL nella vacca da latte
Percorso FAD Aggiornamenti nella gestione dell’allevamento bovino
08-20_FAD_00-00_FAD Ghisleni 26/09/14 15.37 Pagina 18

!!
quando l’utero è tornato in condizione diospitare un embrione. Ciò avviene in gene-re a partire dal 60° giorno dopo il parto. Nel-la fase di NEBAL, è di fondamentale im-portanza quantificare i segnali di bilancioenergetico e proteico negativo per poter met-tere in atto tutte quelle misure necessarienon tanto per impedirne la comparsa quan-to per limitarne ampiezza e gravità. Per da-re un valore alle “dimensioni” del NEBALbasti pensare che il deficit di calorie equi-vale a 1,7 kg al giorno di peso corporeo e pergli amminoacidi di 1,5 kg.
Sistemi di valutazione delNEBAL nella vacca da latte
1. Body condition scoreIl BCS altro non è che la valutazione sog-
gettiva dell’entità delle riserve lipidicheattraverso l’osservazione della quantitàdi grasso sottocutaneo presente sulla grop-pa e sulle apofisi trasverse delle vertebrelombari, ossia al di sopra della fossa delfianco. La scala solitamente utilizzata èquella da 1 a 5, proposta da Edmonsonnel 1988.Tale rilevazione andrebbe fatta alla mes-sa in asciutta e nell’imminenza del par-to, per verificare lo stato di ingrassamentodelle bovine alla fine della lattazione pre-cedente o delle manze, che solitamentevengono mescolate con le vacche adulteproprio a 7 mesi di gravidanza. Inoltre,la doppia valutazione di questa fase per-mette di verificare se ci sono oscillazio-ni, incremento o decremento delle riser-ve lipidiche. Entrambe le condizioni sono sicuramen-te un fattore di rischio per le malattie me-taboliche della fase di transizione e perla fertilità futura delle bovine. Dopo ilparto si esegue una rilevazione a 30-40giorni di lattazione per quantificare l’en-tità delle riserve corporee. Si consideraottimale un punteggio di 3,50 alla messain asciutta e al momento del parto, ac-cettando uno 0,25 in più per le manze.Ideale sarebbe che la perdita di peso do-po il parto rimanesse nell’ambito del mez-zo punto.
19N° 8 Ottobre 2014 ANIMALI DA REDDITO
DOSSIER N. 7 - Alimentazione
NEBALSensibilità/
specificità %Cutt-off N° Inferiore Superiore
al 10% al 25%Grasso % > 4,8 771 39/87 28/89Proteina % < 2,9 771 17/85 18/86Lattosio % < 4,5 771 27/87 22/88Rapporto grasso/proteine > 1,4 771 66/68 61/73
> 1,5 771 51/87 41/91> 1,6 771 29/96 19/98
Ketolac µmol/l > 100 726 29/91 24/93> 200 726 14/98 10/99
Nitroprusside Doubtful+pos 733 31/94 24/97Cleary pos 733 18/98 12/99
BCS ≥ 3,5 759 23/77 23/77BHBA mmol/l > 1,0 376 28/93 19/96
> 1,2 376 25/97 14/98
TABELLA 4. Sensibilità e specificità dei test di latte, bcs, corpi chetonici nel latte e metaboliti ematici
per diagnosticare il bilancio energetico negativo
Vacche (72) nel periodo tra 2 e 12 settimane post partum misuratesettimanalmente per 9-11 volte
Parametro r PMetaboliti ematiciNEFA - 0,685 < 0,001Glucosio 0,456 < 0,001BHBA - 0,451 < 0,001Colesterolo 0,406 < 0,001Creatinina - 0,415 < 0,001Urea 0,259 < 0,001Albumina 0,137 < 0,001Ormoni EmaticiTiroxina 0,418 < 0,001IGF-1 0,320 < 0,0013,5,3-triodotironina 0,270 < 0,001Insulina 0,230 < 0,001GH - 0,217 < 0,001Leptina - 0,027 0,437Enzimi ematiciLattato deidrogenasi - 0,199 < 0,001Aspartato amino transferasi - 0,154 < 0,001Glutammato deidrogenasi - 0,114 < 0,001LatteRapporto grasso:lattosio - 0,589 < 0,001Grasso - 0,565 < 0,001Rapporto grasso:proteina - 0,496 < 0,001Acetone - 0,410 < 0,001Lattosio 0,363 < 0,001Proteina - 0,185 < 0,001Urea 0,103 0,002
TABELLA 3. Correlazionedi Pearson di parametri ematici
e del latte con il bilancioenergetico tra la prima e la decima settimana
dopo il parto
08-20_FAD_00-00_FAD Ghisleni 26/09/14 15.37 Pagina 19

2. BiomarkersEsistono molti parametri ematici, comemetaboliti ed enzimi, o parametri del lat-te che possono e potrebbero essere uti-lizzati con precisione per valutare l’am-piezza del bilancio energetico e proteiconegativo (tabella 3).Tra i metaboliti ematici sicuramente i piùimportanti sono i NEFA, ossia acidi gras-si che vengono liberati dal tessuto adi-poso in condizioni di NEBAL. È consi-derata come valore soglia, prima del par-to, una concentrazione ematica di NEFAdi 0,29 mmol o meq/l, mentre dopo il par-to di 0,6. Un altro parametro ematico disicuro interesse è l’IGF-1. Questo ormo-ne, la cui secrezione epatica dipende dalGH, ma anche dall’insulina e dalla con-centrazione ematica degli amminoacidi,è il più potente fattore endocrino di cre-scita follicolare oggi conosciuto.Di grande interesse pratico ed economi-co ai fini della valutazione del bilancioenergetico e proteico negativo sono al-cuni parametri del latte individuale (nondi massa!), come la percentuale di gras-so, proteine e lattosio. Dalle molte ricer-che effettuate è stato osservato che la per-centuale di grasso del latte a inizio latta-zione (0-75 giorni) può essere indicativodi un forte dimagrimento della bovina, inonore del principio che avendo la mam-mella la priorità metabolica, in questa fa-se recluta gli acidi grassi ematici per lasintesi del grasso del latte. Lo stesso puòdirsi se la percentuale di proteina nel lat-te, nel medesimo periodo, scende al disotto del 2,9%, a indicare un bilancio ener-getico negativo, ma soprattutto ammi-noacidico. Di più facile utilizzo pratico èil rapporto grasso/proteine. Valori > 1,4e > 1,5 possono essere considerati bio-markers utilizzabili. Tra questi rapportitra il grasso e le proteine del latte e il bi-
lancio energetico esiste una correlazionenegativa (0,36-0,74) (tabella 4).Con le dovute cautele, si può utilizzare laconcentrazione di lattosio nel latte indi-viduale nelle prime settimane di latta-zione solo nelle bovine senza infiamma-zioni delle mammella quantificabili conuna concentrazione di cellule somatiche> 200.000 ml. Un livello di lattosio < 4,5%può aiutare a diagnosticare un grave NE-BAL. Tutti questi parametri del latte so-no stati adottati dall’Associazione italia-na allevatori in un due report, detti “Sin-tetico collettivo e individuale”, che ven-gono restituiti all’allevatore che parteci-pa ai controlli funzionali dopo il controlloperiodico delle produzioni del latte e deisuoi costituenti.Lo sviluppo delle tecnologie real time ap-plicabili alle sale di mungitura renderàdisponibili in futuro molti altri biomar-kers. ■
20 N° 8 Ottobre 2014ANIMALI DA REDDITO
Per saperne di più 1-Baldwin R.L. et al. Manipulating metabolic para-meters to improve growth rate and milk secretion.J. Animal. Sci., 1980; vol. 51, n. 6: pp. 1416-1428.2-Coppock C.E. Energy nutrition and metabolism oflactating dairy cows. J. Dairy Sci., 1985; vol. 68: pp.3403-3410.3-Greenfield R.B. Change in mRNA expression forgluconeogenic enzymes in liver of dairy cattle du-ring the transition to lactation. J. Dairy Sci., 2000;vol. 83: pp. 1228-1236.4-Lemosquet S. et al. Effects of glucose, propionicacid, and nonessential amino acids on glucose me-tabolism and milk yield in Holstein dairy cows. J.Dairy Sci., 2009; vol. 92: pp. 3244-3257.5-Moe P.W. Energy metabolism of dairy cattle. J.Dairy Sci., 1981; vol. 64: pp. 1120-1139.6-National Research Council. Nutrient requirementsof dairy cattle, 2001; pp.13-257-Weiss W.P. Refining the net energy system. WCDSAdvances in dairy technology, 2010; vol. 22: pp.191-202.8-Weiss W.P. Estimating the available energy con-tenent of feed for dairy cattle. J. Dairy Sci., 1998;vol. 81: pp. 830-839.
Percorso FAD Aggiornamenti nella gestione dell’allevamento bovino
08-20_FAD_00-00_FAD Ghisleni 26/09/14 15.37 Pagina 20


















